

Gut microbiota derived metabolites contribute to intestinal barrier maturation at the suckling-to-weaning transition
- PMID: 32352849
- PMCID: PMC7524271
- DOI: 10.1080/19490976.2020.1747335
Gut microbiota derived metabolites contribute to intestinal barrier maturation at the suckling-to-weaning transition
Abstract
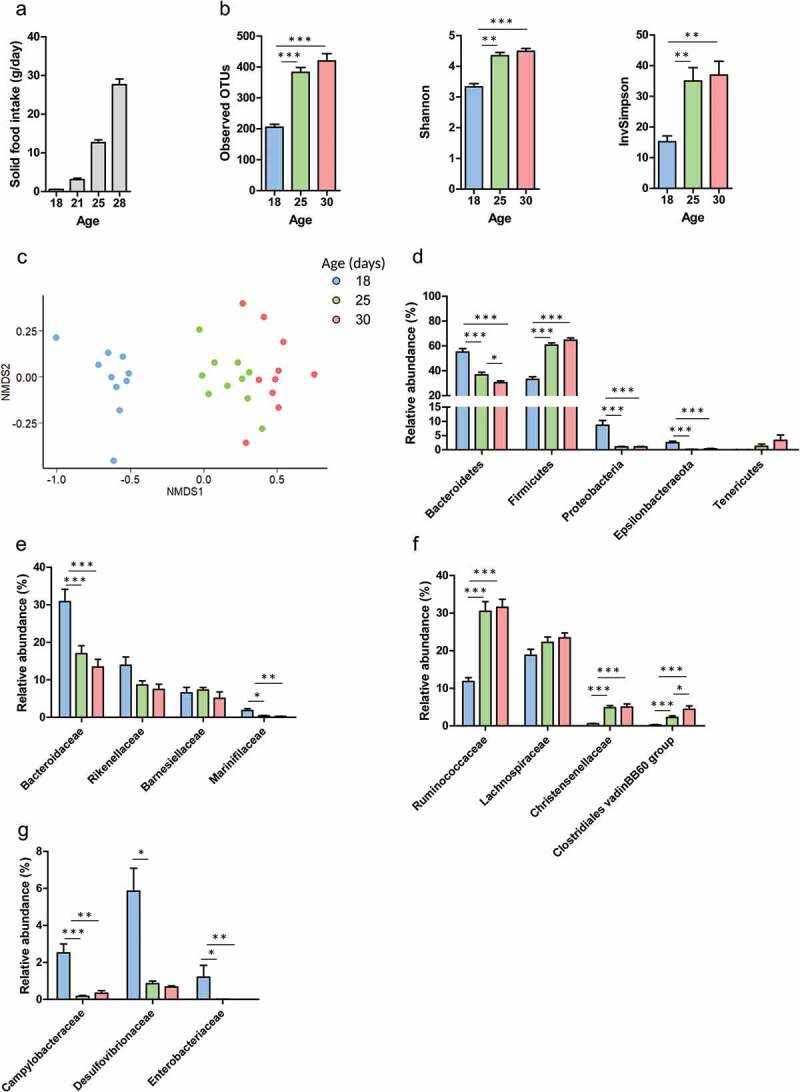
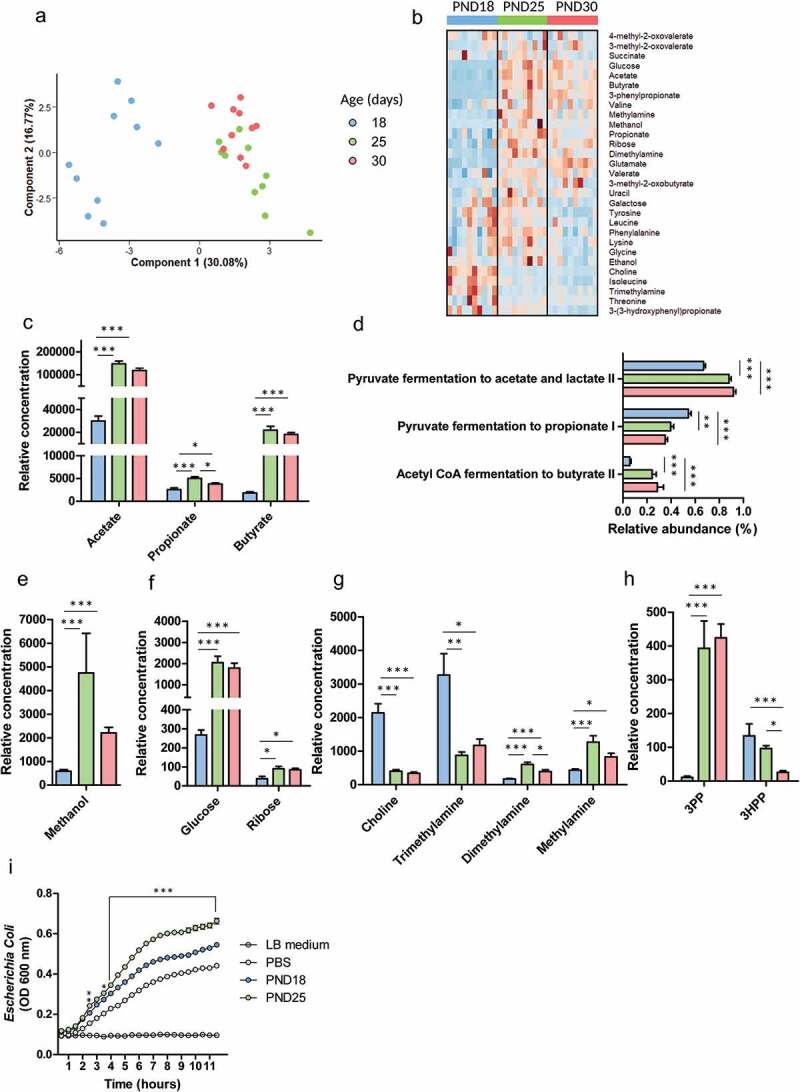
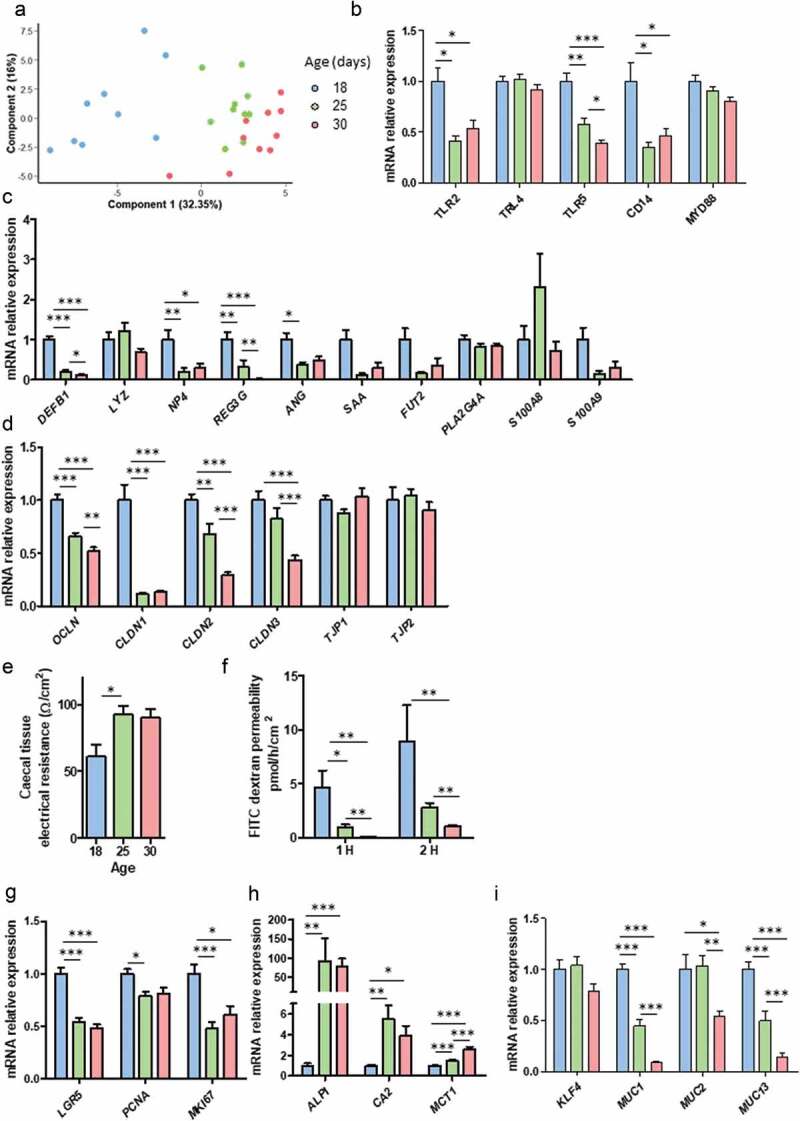
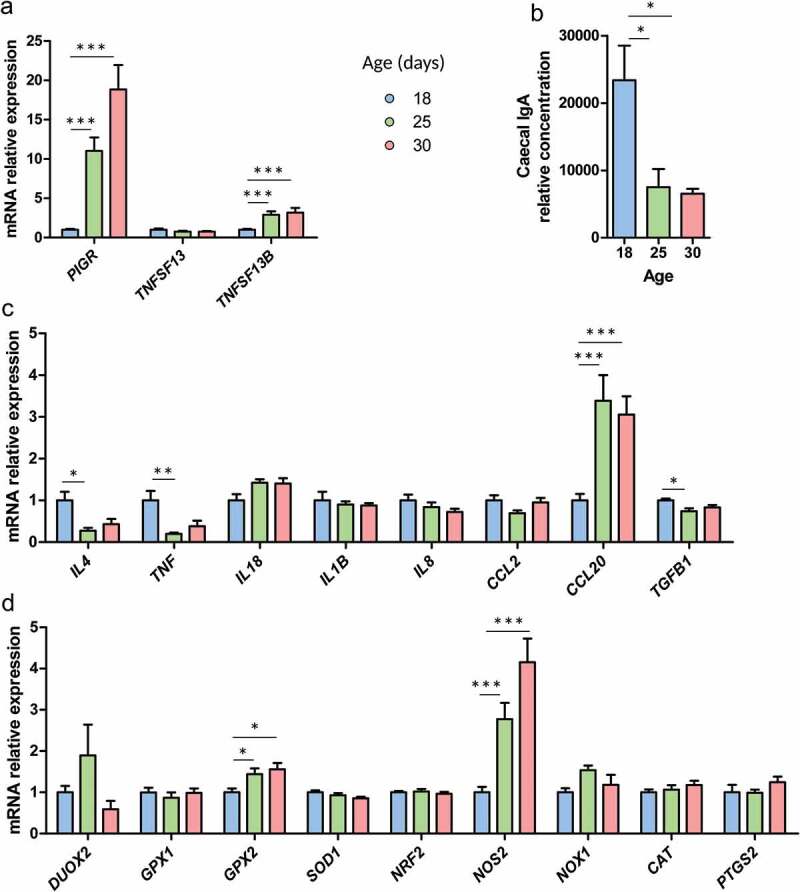
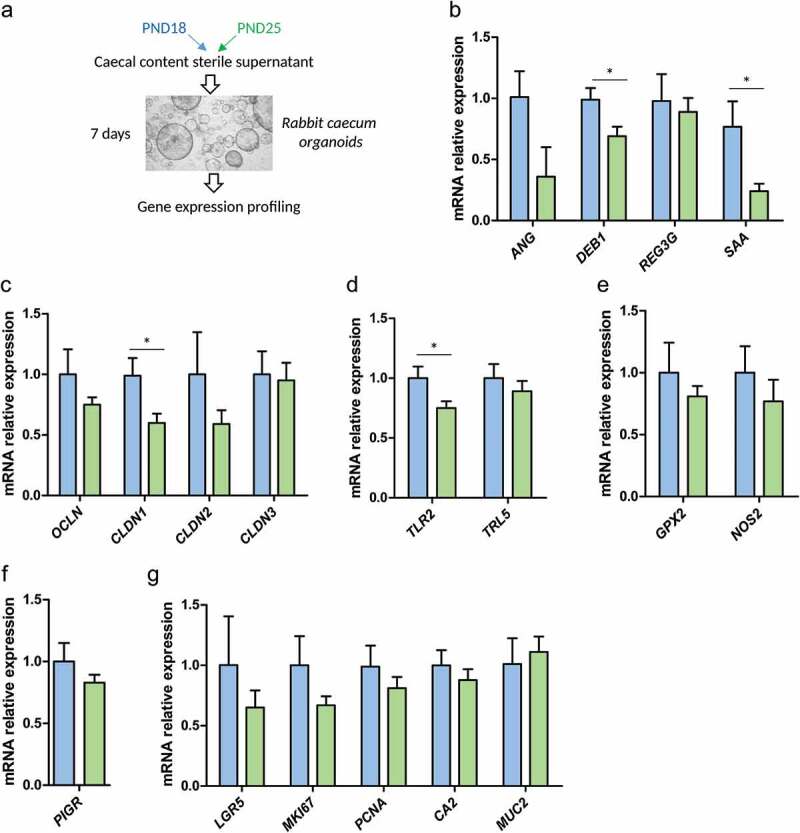
In suckling mammals, the onset of solid food ingestion is coincident with the maturation of the gut barrier. This ontogenic process is driven by the colonization of the intestine by the microbiota. However, the mechanisms underlying the microbial regulation of the intestinal development in early life are not fully understood. Here, we studied the co-maturation of the microbiota (composition and metabolic activity) and of the gut barrier at the suckling-to-weaning transition by using a combination of experiments in vivo (suckling rabbit model), ex vivo (Ussing chambers) and in vitro (epithelial cell lines and organoids). The microbiota composition, its metabolic activity, para-cellular epithelial permeability and the gene expression of key components of the gut barrier shifted sharply at the onset of solid food ingestion in vivo, despite milk was still predominant in the diet at that time. We found that cecal content sterile supernatant (i.e. containing a mixture of metabolites) obtained after the onset of solid food ingestion accelerated the formation of the epithelial barrier in Caco-2 cells in vitro and our results suggested that these effects were driven by the bacterial metabolite butyrate. Moreover, the treatment of organoids with cecal content sterile supernatant partially replicated in vitro the effects of solid food ingestion on the epithelial barrier in vivo. Altogether, our results show that the metabolites produced by the microbiota at the onset of solid food ingestion contribute to the maturation of the gut barrier at the suckling-to-weaning transition. Targeting the gut microbiota metabolic activity during this key developmental window might therefore be a promising strategy to promote intestinal homeostasis.
Keywords: Early life; epithelium; intestinal barrier development; maternal milk; metabolome; organoids; solid food.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources