

IP3-Dependent Ca2+ Oscillations Switch into a Dual Oscillator Mechanism in the Presence of PLC-Linked Hormones
- PMID: 32353764
- PMCID: PMC7191650
- DOI: 10.1016/j.isci.2020.101062
IP3-Dependent Ca2+ Oscillations Switch into a Dual Oscillator Mechanism in the Presence of PLC-Linked Hormones
Abstract
Ca2+ oscillations that depend on inositol-1,4,5-trisphosphate (IP3) have been ascribed to biphasic Ca2+ regulation of the IP3 receptor (IP3R) or feedback mechanisms controlling IP3 levels in different cell types. IP3 uncaging in hepatocytes elicits Ca2+ transients that are often localized at the subcellular level and increase in magnitude with stimulus strength. However, this does not reproduce the broad baseline-separated global Ca2+ oscillations elicited by vasopressin. Addition of hormone to cells activated by IP3 uncaging initiates a qualitative transition from high-frequency spatially disorganized Ca2+ transients, to low-frequency, oscillatory Ca2+ waves that propagate throughout the cell. A mathematical model with dual coupled oscillators that integrates Ca2+-induced Ca2+ release at the IP3R and mutual feedback mechanisms of cross-coupling between Ca2+ and IP3 reproduces this behavior. Thus, multiple Ca2+ oscillation modes can coexist in the same cell, and hormonal stimulation can switch from the simpler to the more complex to yield robust signaling.
Keywords: Cell Biology; Mathematical Biosciences; Specialized Functions of Cells.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures


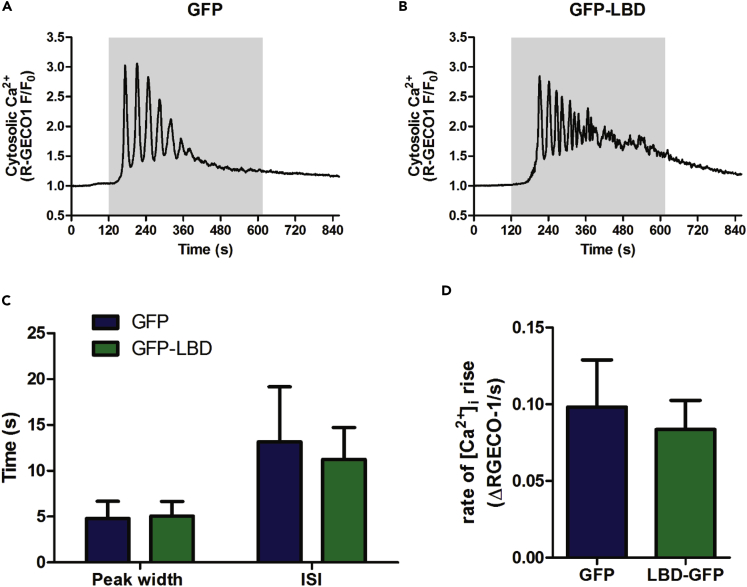


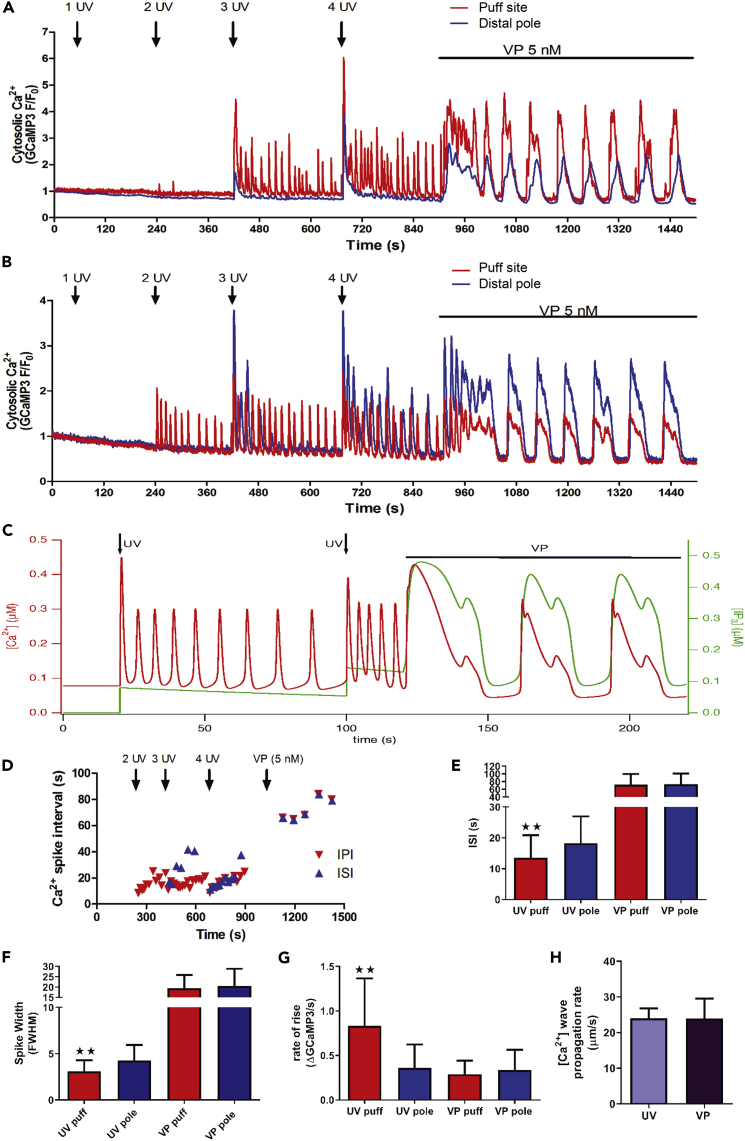
References
-
- Arguin G., Regimbald-Dumas Y., Fregeau M.-O., Caron A.Z., Guillemette G. Protein kinase C phosphorylates the inositol 1,4,5-trisphosphate receptor type 2 and decreases the mobilization of Ca2+in pancreatoma AR4-2J cells. J. Endocrinol. 2007;192:659–668. - PubMed
-
- Baffy G., Yang L., Michalopoulos G.K., Williamson J.R. Hepatocyte growth factor induces calcium mobilization and inositol phosphate production in rat hepatocytes. J. Cell Physiol. 1992;153:332–339. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
