

Gut Microbiota-Derived Short-Chain Fatty Acids Promote Poststroke Recovery in Aged Mice
- PMID: 32354259
- PMCID: PMC7415518
- DOI: 10.1161/CIRCRESAHA.119.316448
Gut Microbiota-Derived Short-Chain Fatty Acids Promote Poststroke Recovery in Aged Mice
Abstract
Rationale: The elderly experience profound systemic responses after stroke, which contribute to higher mortality and more severe long-term disability. Recent studies have revealed that stroke outcomes can be influenced by the composition of gut microbiome. However, the potential benefits of manipulating the gut microbiome after injury is unknown.
Objective: To determine if restoring youthful gut microbiota after stroke aids in recovery in aged subjects, we altered the gut microbiome through young fecal transplant gavage in aged mice after experimental stroke. Further, the effect of direct enrichment of selective bacteria producing short-chain fatty acids (SCFAs) was tested as a more targeted and refined microbiome therapy.
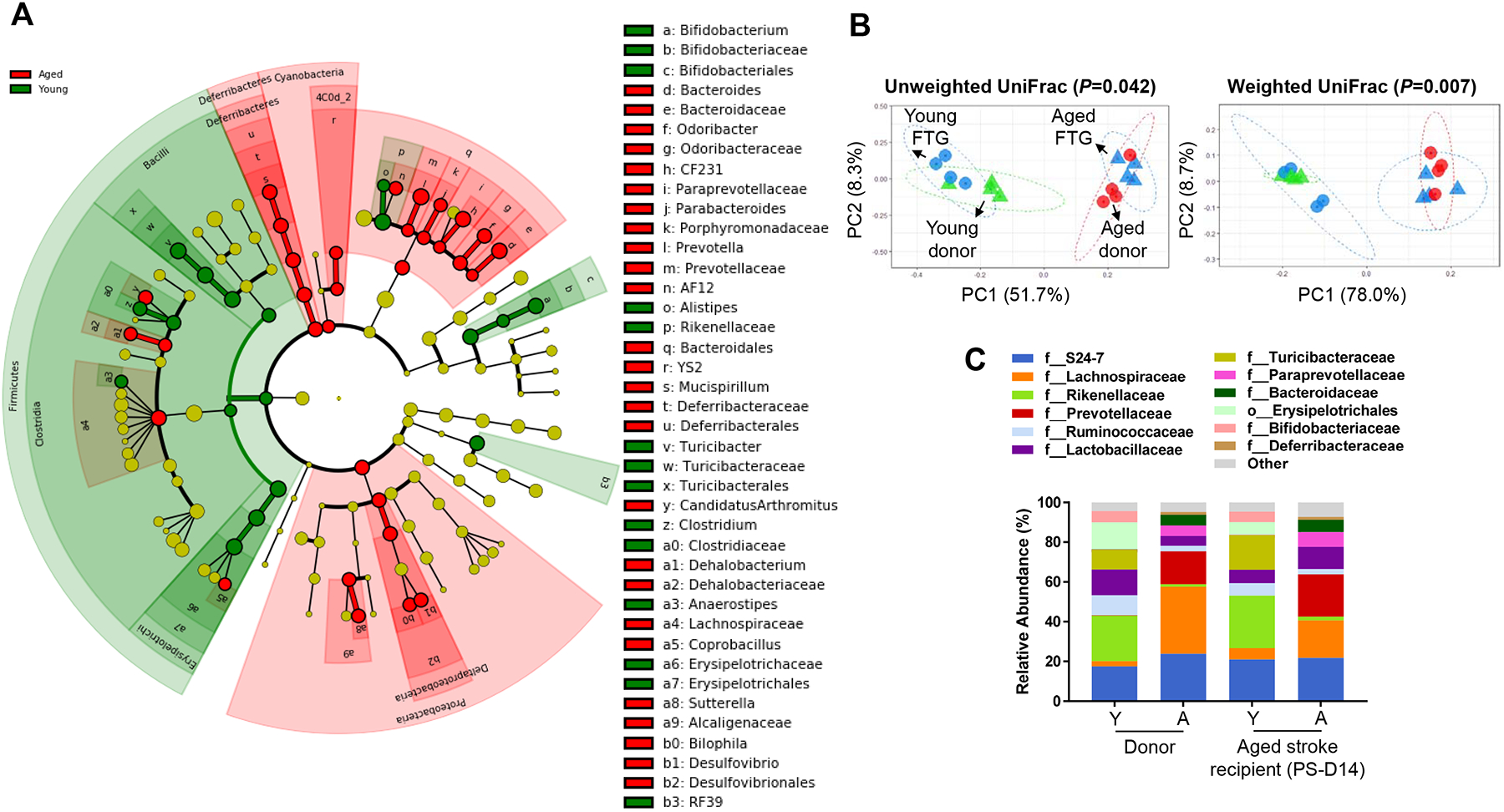
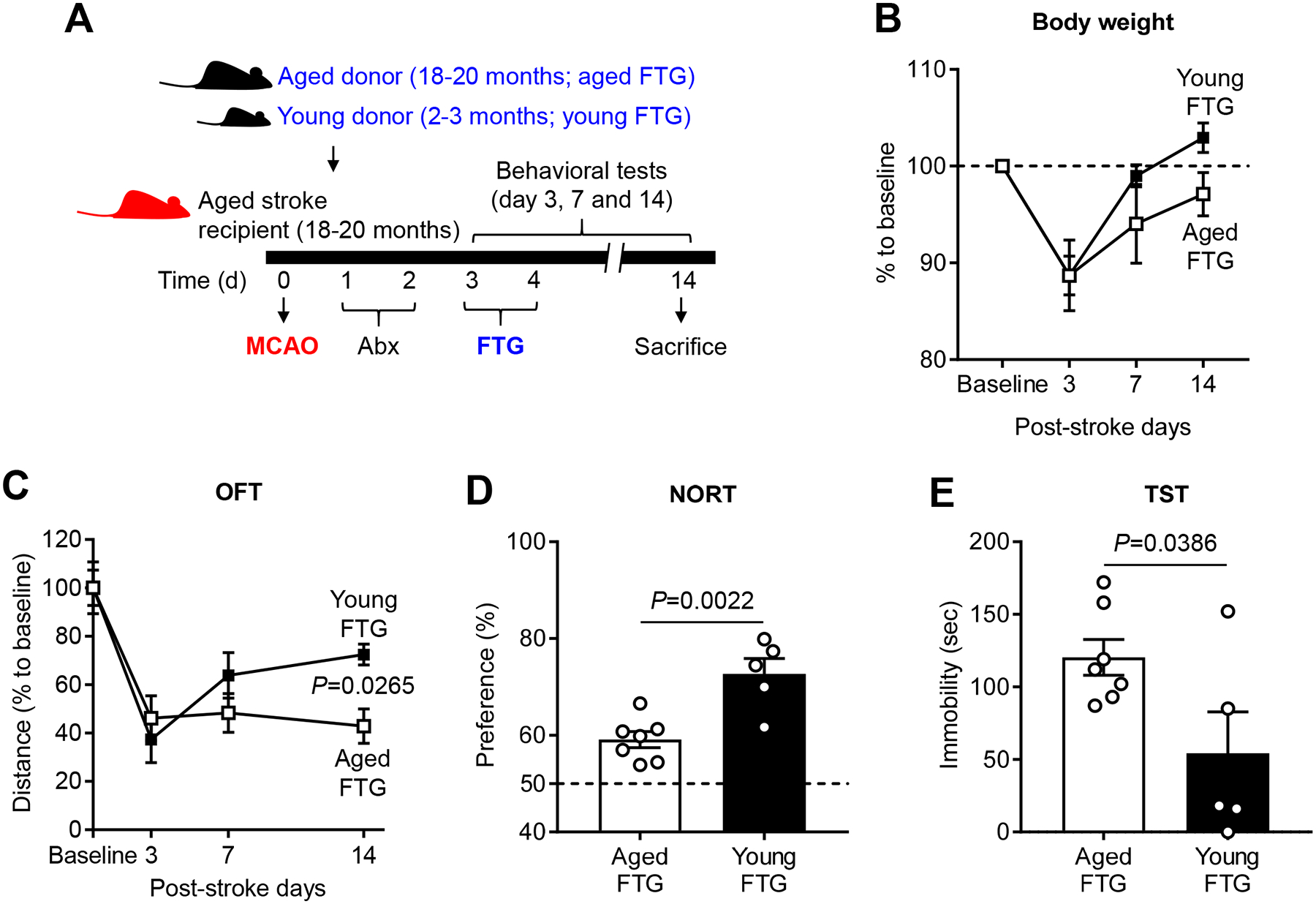
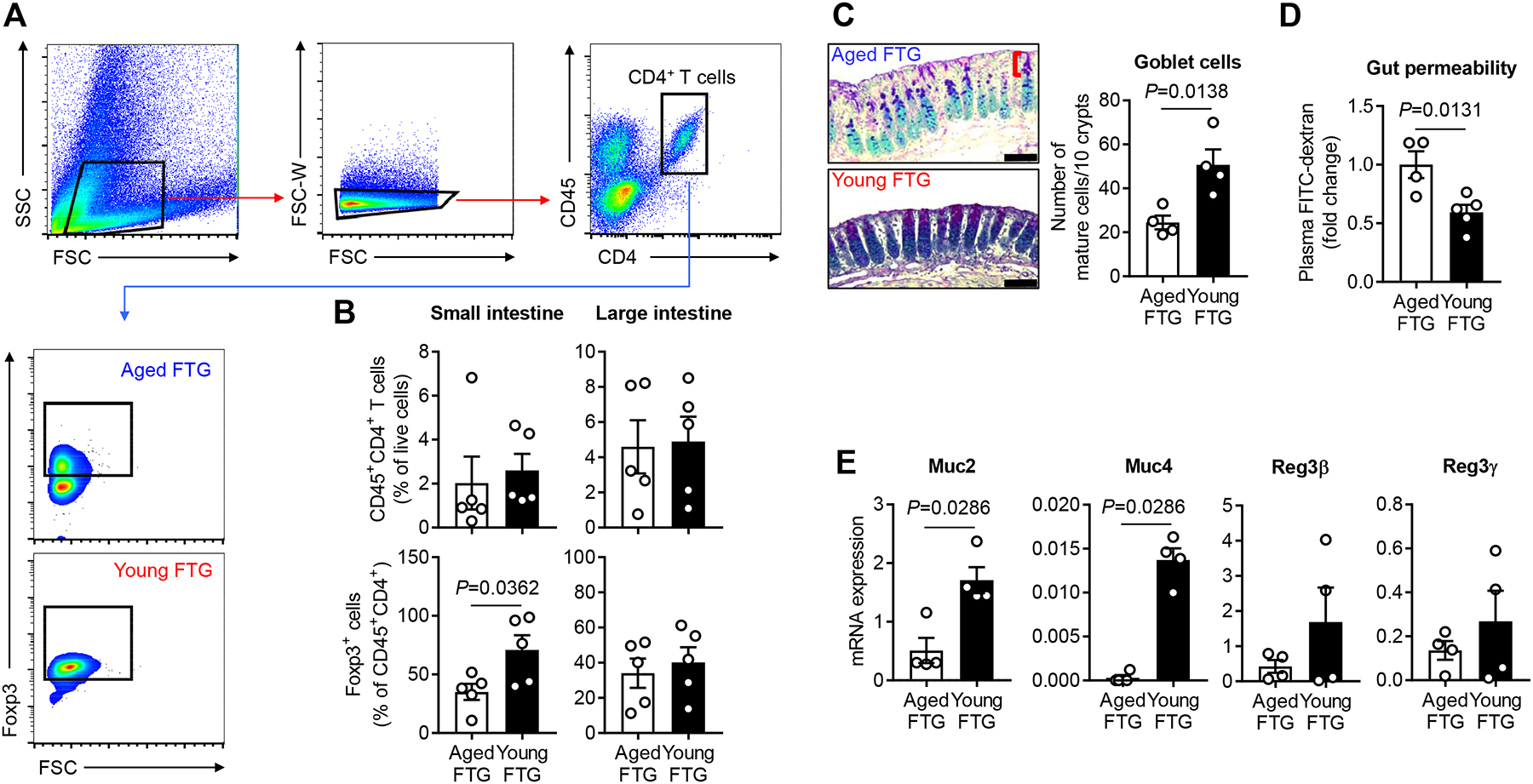
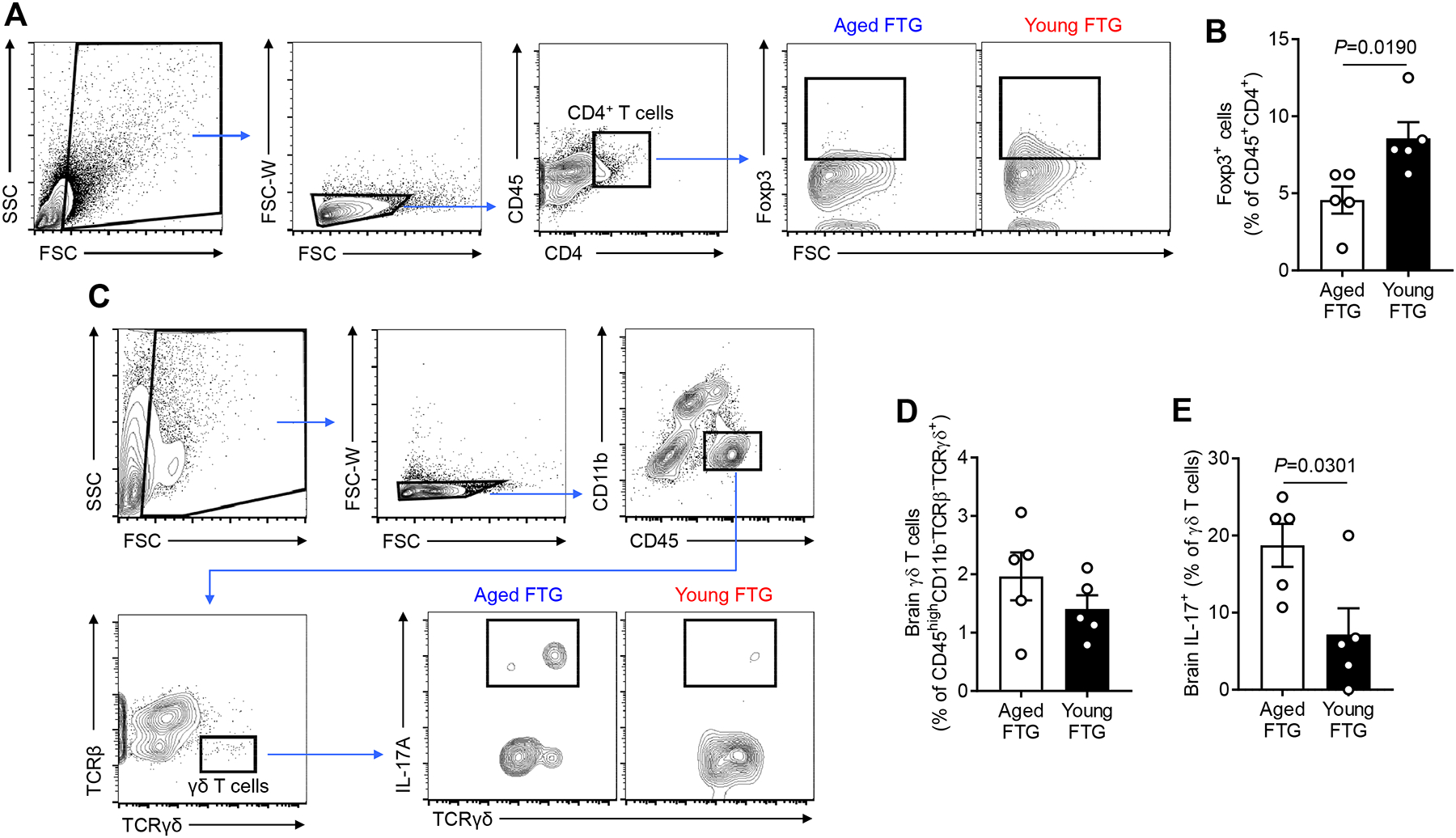
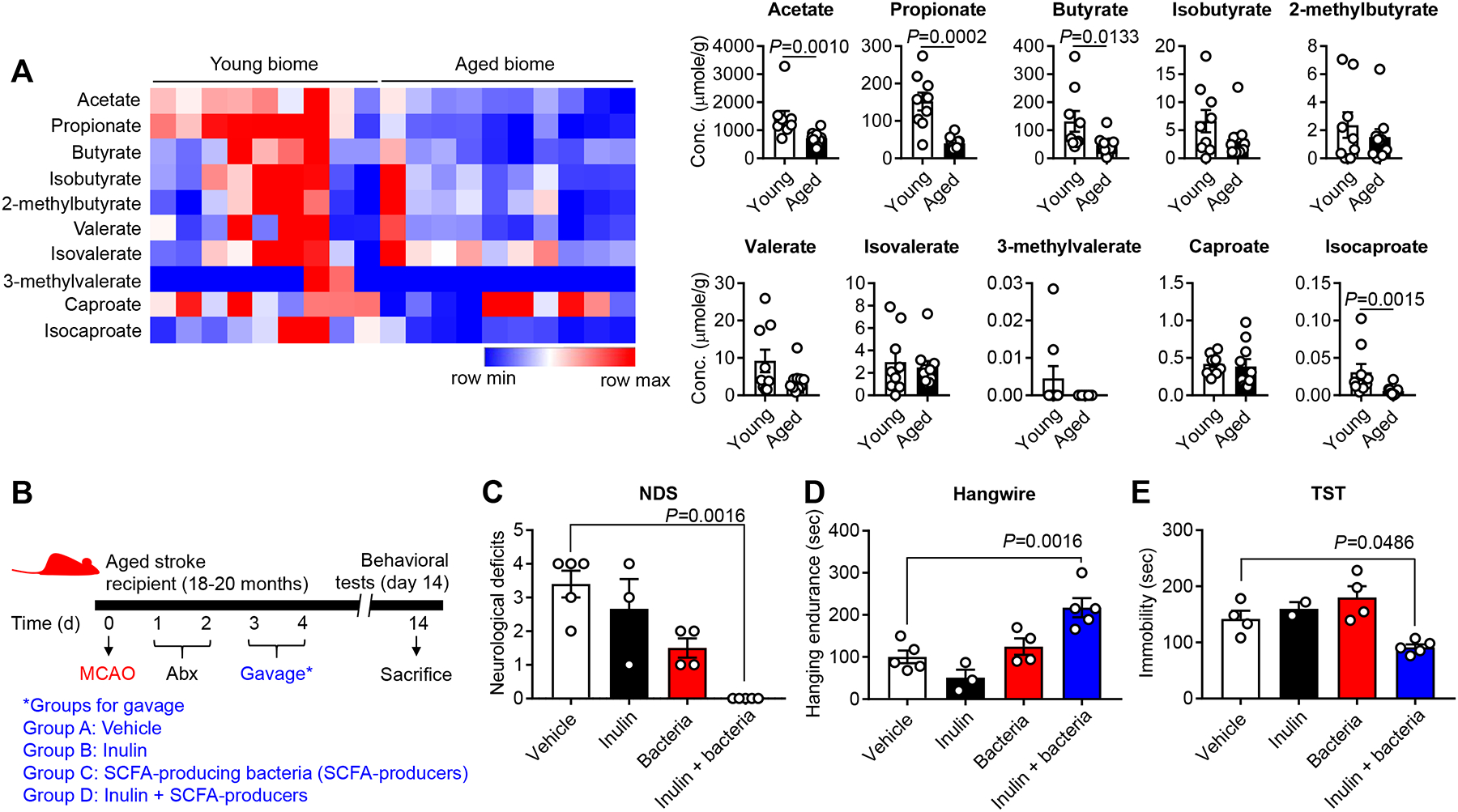
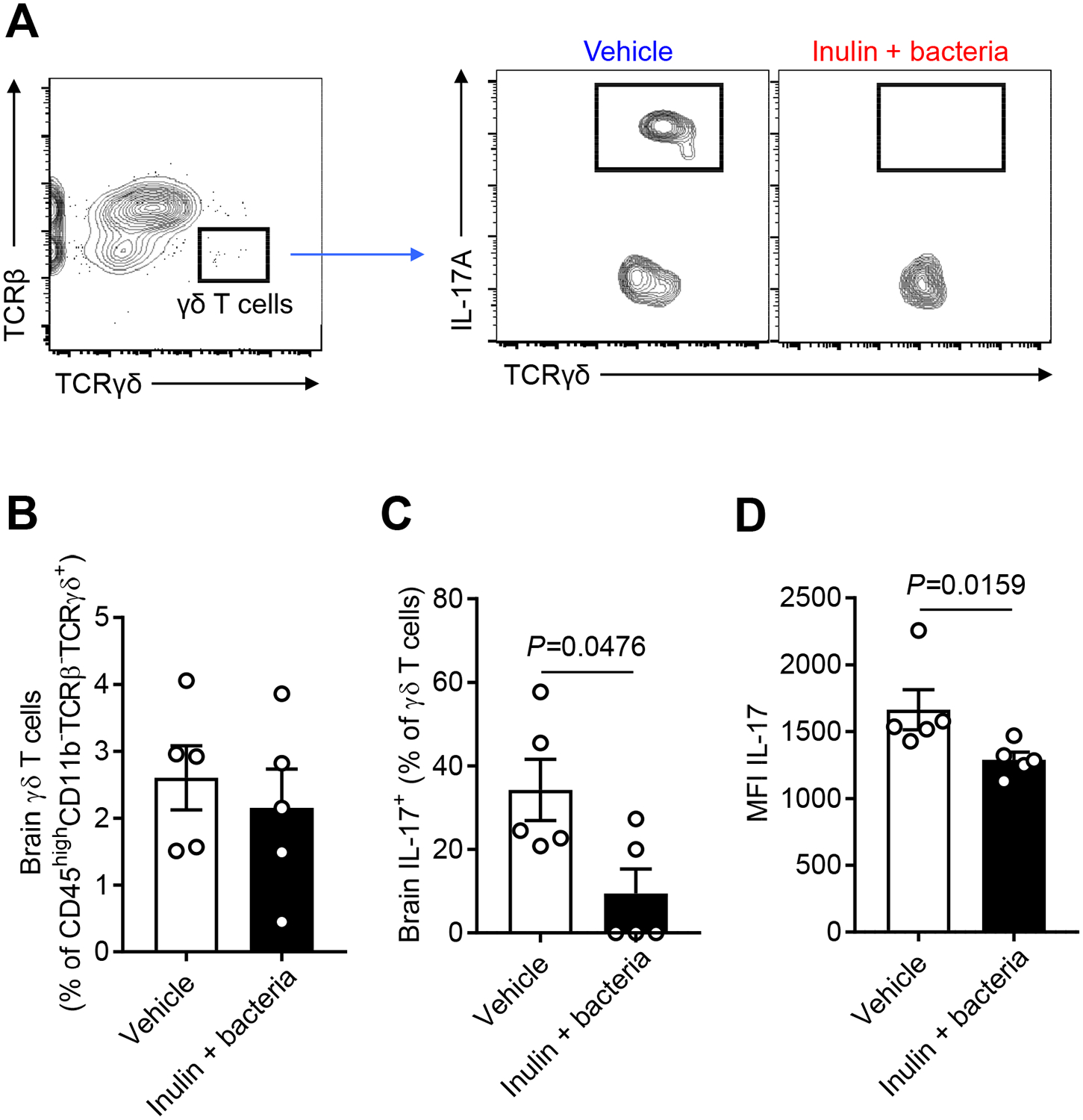
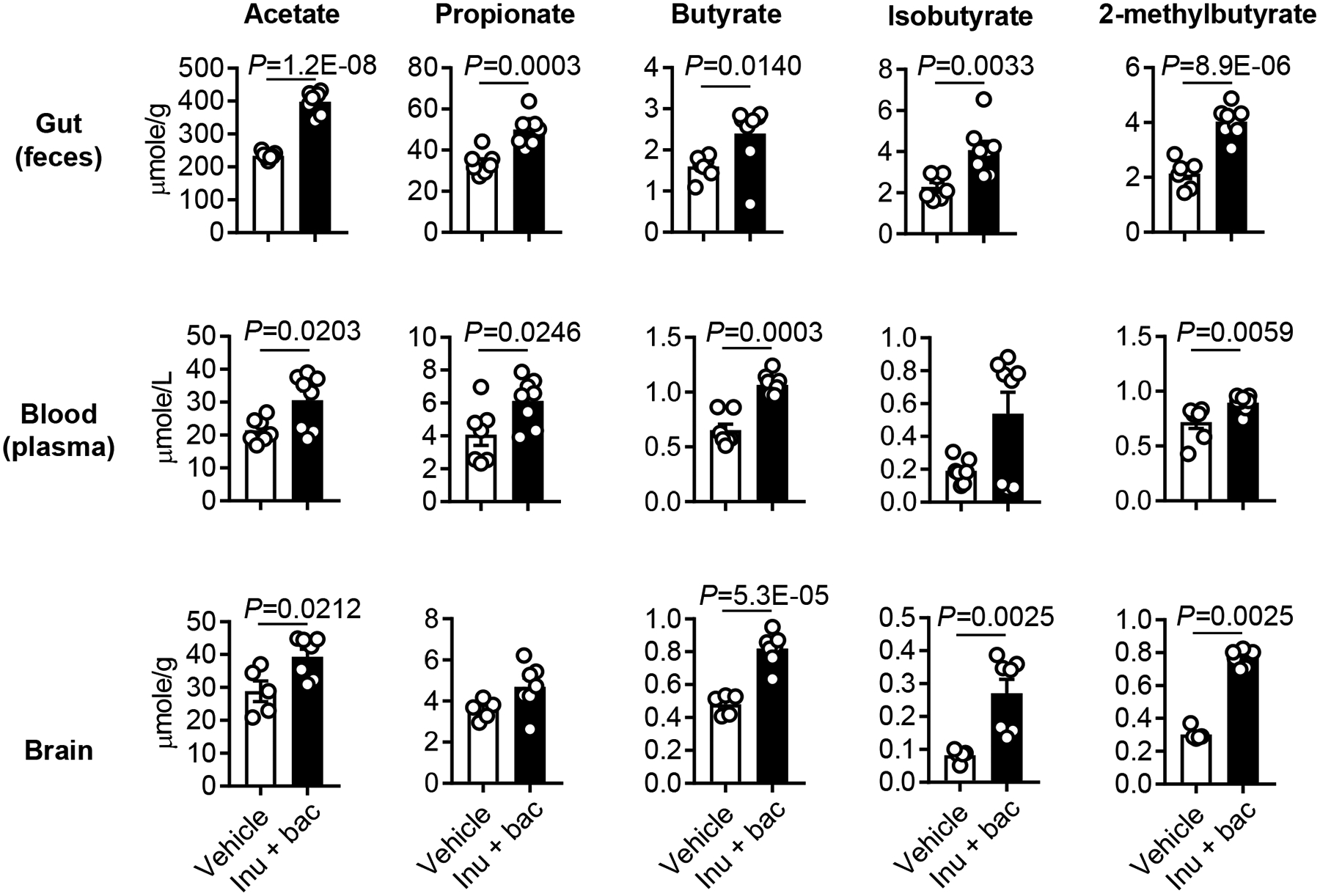
Methods and results: Aged male mice (18-20 months) were subjected to ischemic stroke by middle cerebral artery occlusion. We performed fecal transplant gavage 3 days after middle cerebral artery occlusion using young donor biome (2-3 months) or aged biome (18-20 months). At day 14 after stroke, aged stroke mice receiving young fecal transplant gavage had less behavioral impairment, and reduced brain and gut inflammation. Based on data from microbial sequencing and metabolomics analysis demonstrating that young fecal transplants contained much higher SCFA levels and related bacterial strains, we selected 4 SCFA-producers (Bifidobacterium longum, Clostridium symbiosum, Faecalibacterium prausnitzii, and Lactobacillus fermentum) for transplantation. These SCFA-producers alleviated poststroke neurological deficits and inflammation, and elevated gut, brain and plasma SCFA concentrations in aged stroke mice.
Conclusions: This is the first study suggesting that the poor stroke recovery in aged mice can be reversed via poststroke bacteriotherapy following the replenishment of youthful gut microbiome via modulation of immunologic, microbial, and metabolomic profiles in the host.
Keywords: aging; fecal microbiota transplantation; gut microbiota; inflammation; metabolomics; middle cerebral artery occlusion.
Figures
References
-
- Stanley D, Mason LJ, Mackin KE, Srikhanta YN, Lyras D, Prakash MD, Nurgali K, Venegas A, Hill MD, Moore RJ, Wong CHY. Translocation and dissemination of commensal bacteria in post-stroke infection. Nature Medicine. 2016;22:1277 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
