

CLASP Mediates Microtubule Repair by Restricting Lattice Damage and Regulating Tubulin Incorporation
- PMID: 32359430
- PMCID: PMC7280784
- DOI: 10.1016/j.cub.2020.03.070
CLASP Mediates Microtubule Repair by Restricting Lattice Damage and Regulating Tubulin Incorporation
Abstract
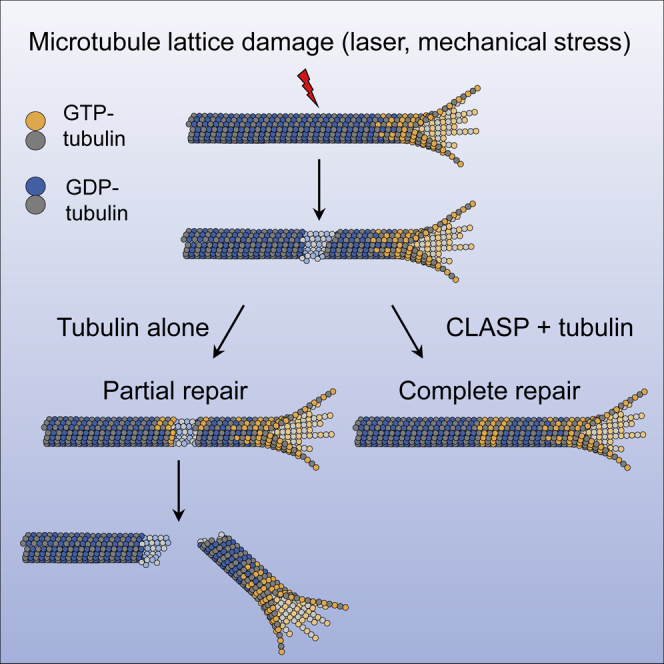
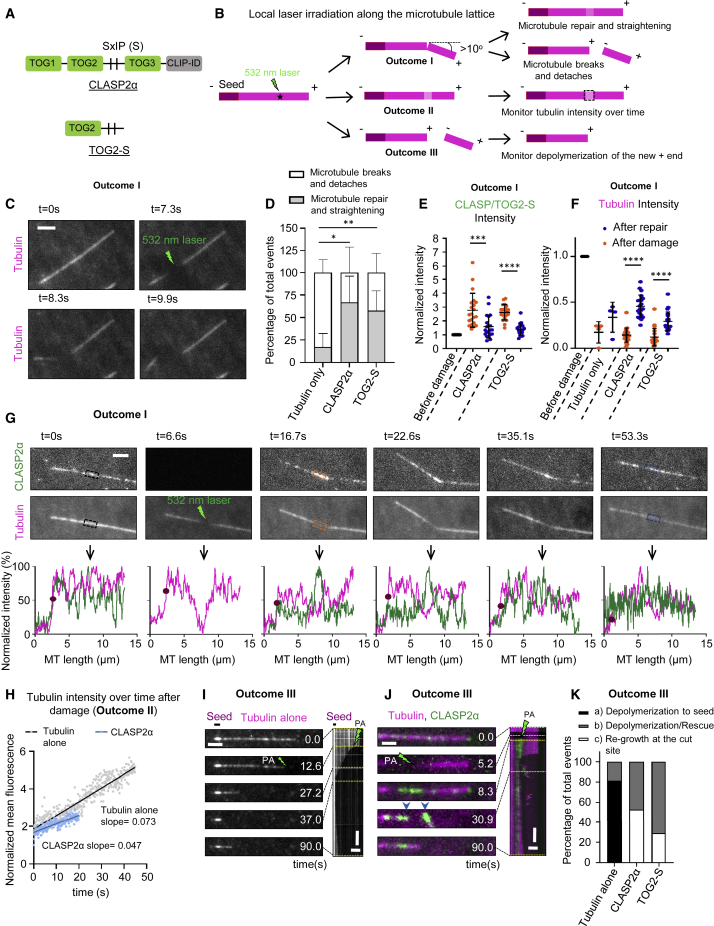
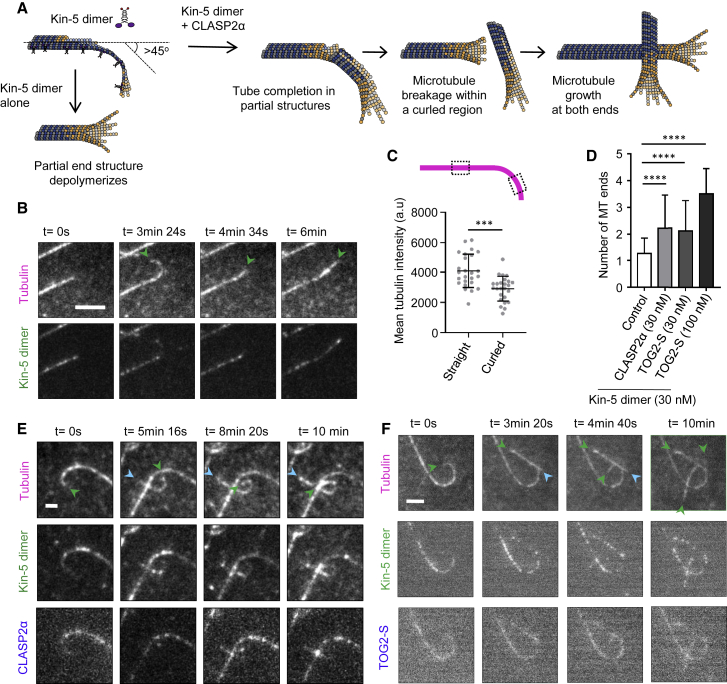
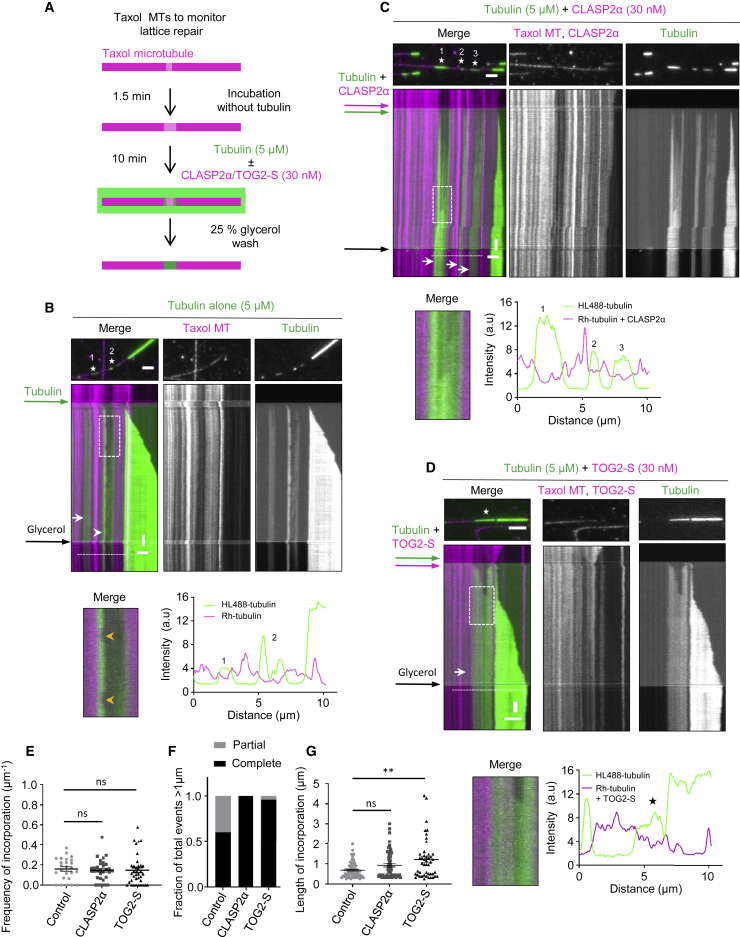
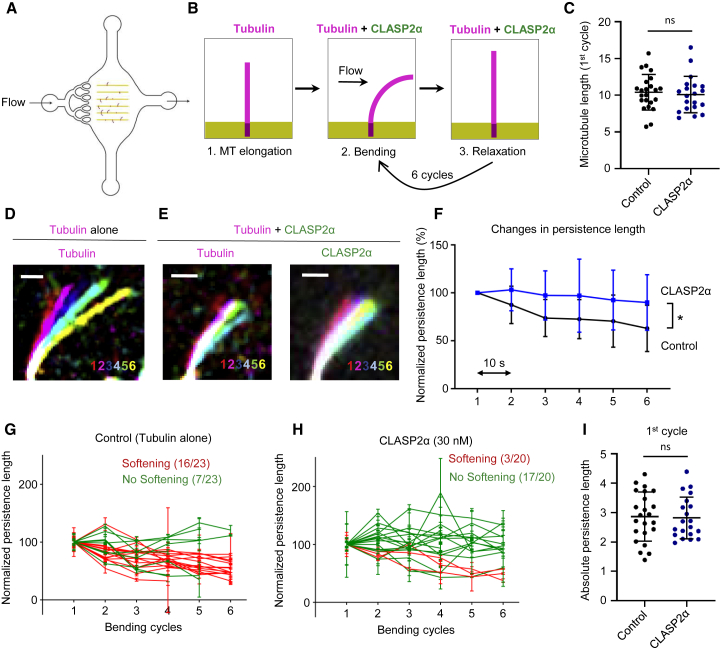
Microtubules play a key role in cell division, motility, and intracellular trafficking. Microtubule lattices are generally regarded as stable structures that undergo turnover through dynamic instability of their ends [1]. However, recent evidence suggests that microtubules also exchange tubulin dimers at the sites of lattice defects, which can be induced by mechanical stress, severing enzymes, or occur spontaneously during polymerization [2-6]. Tubulin incorporation can restore microtubule integrity; moreover, "islands" of freshly incorporated GTP-tubulin can inhibit microtubule disassembly and promote rescues [3, 4, 6-8]. Microtubule repair occurs in vitro in the presence of tubulin alone [2-6, 9]. However, in cells, it is likely to be regulated by specific factors, the nature of which is currently unknown. CLASPs are interesting candidates for microtubule repair because they induce microtubule nucleation, stimulate rescue, and suppress catastrophes by stabilizing incomplete growing plus ends with lagging protofilaments and promoting their conversion into complete ones [10-17]. Here, we used in vitro reconstitution assays combined with laser microsurgery and microfluidics to show that CLASP2α indeed stimulates microtubule lattice repair. CLASP2α promoted tubulin incorporation into damaged lattice sites, thereby restoring microtubule integrity. Furthermore, it induced the formation of complete tubes from partial protofilament assemblies and inhibited microtubule softening caused by hydrodynamic-flow-induced bending. The catastrophe-suppressing domain of CLASP2α, TOG2, combined with a microtubule-tethering region, was sufficient to stimulate microtubule repair, suggesting that catastrophe suppression and lattice repair are mechanistically similar. Our results suggest that the cellular machinery controlling microtubule nucleation and growth can also help to maintain microtubule integrity.
Keywords: CLASP; TOG2; in vitro reconstitution; laser microsurgery; lattice defects; microtubule catastrophe; microtubule dynamics; microtubule repair; microtubule rescue; tubulin.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
Comment in
-
Cytoskeletal Repair: Microtubule Orthopaedics to the Rescue.Curr Biol. 2020 Jun 8;30(11):R646-R649. doi: 10.1016/j.cub.2020.04.049. Curr Biol. 2020. PMID: 32516614
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
