

Understanding the interplay between CpG island-associated gene promoters and H3K4 methylation
- PMID: 32360393
- PMCID: PMC7294231
- DOI: 10.1016/j.bbagrm.2020.194567
Understanding the interplay between CpG island-associated gene promoters and H3K4 methylation
Abstract
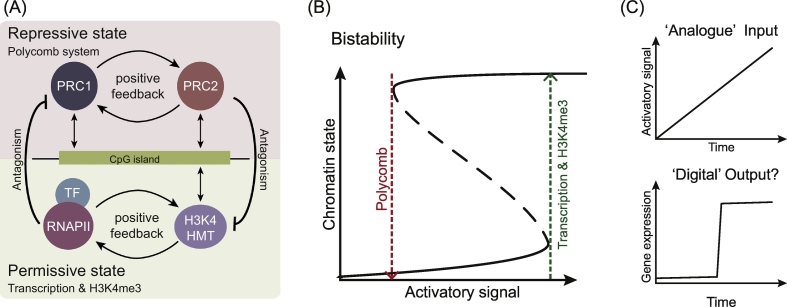
The precise regulation of gene transcription is required to establish and maintain cell type-specific gene expression programs during multicellular development. In addition to transcription factors, chromatin, and its chemical modification, play a central role in regulating gene expression. In vertebrates, DNA is pervasively methylated at CG dinucleotides, a modification that is repressive to transcription. However, approximately 70% of vertebrate gene promoters are associated with DNA elements called CpG islands (CGIs) that are refractory to DNA methylation. CGIs integrate the activity of a range of chromatin-regulating factors that can post-translationally modify histones and modulate gene expression. This is exemplified by the trimethylation of histone H3 at lysine 4 (H3K4me3), which is enriched at CGI-associated gene promoters and correlates with transcriptional activity. Through studying H3K4me3 at CGIs it has become clear that CGIs shape the distribution of H3K4me3 and, in turn, H3K4me3 influences the chromatin landscape at CGIs. Here we will discuss our understanding of the emerging relationship between CGIs, H3K4me3, and gene expression.
Keywords: Chromatin; CpG islands; DNA methylation; H3K4me3; Transcription.
Copyright © 2020 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
