

Chloroplast-to-nucleus retrograde signalling controls intercellular trafficking via plasmodesmata formation
- PMID: 32362251
- PMCID: PMC7209952
- DOI: 10.1098/rstb.2019.0408
Chloroplast-to-nucleus retrograde signalling controls intercellular trafficking via plasmodesmata formation
Abstract
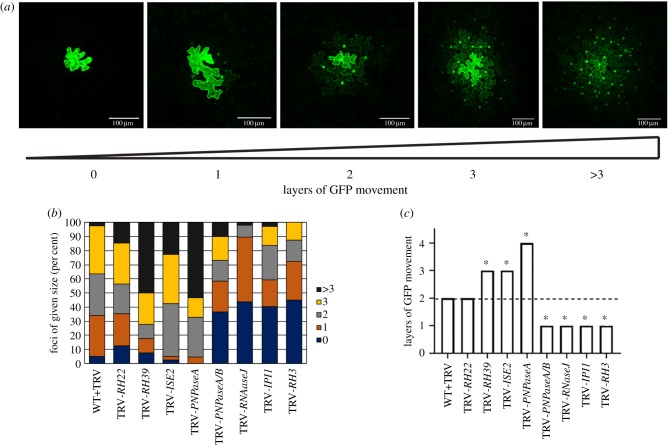
The signalling pathways that regulate intercellular trafficking via plasmodesmata (PD) remain largely unknown. Analyses of mutants with defects in intercellular trafficking led to the hypothesis that chloroplasts are important for controlling PD, probably by retrograde signalling to the nucleus to regulate expression of genes that influence PD formation and function, an idea encapsulated in the organelle-nucleus-PD signalling (ONPS) hypothesis. ONPS is supported by findings that point to chloroplast redox state as also modulating PD. Here, we have attempted to further elucidate details of ONPS. Through reverse genetics, expression of select nucleus-encoded genes with known or predicted roles in chloroplast gene expression was knocked down, and the effects on intercellular trafficking were then assessed. Silencing most genes resulted in chlorosis, and the expression of several photosynthesis and tetrapyrrole biosynthesis associated nuclear genes was repressed in all silenced plants. PD-mediated intercellular trafficking was changed in the silenced plants, consistent with predictions of the ONPS hypothesis. One striking observation, best exemplified by silencing the PNPase homologues, was that the degree of chlorosis of silenced leaves was not correlated with the capacity for intercellular trafficking. Finally, we measured the distribution of PD in silenced leaves and found that intercellular trafficking was positively correlated with the numbers of PD. Together, these results not only provide further support for ONPS but also point to a genetic mechanism for PD formation, clarifying a longstanding question about PD and intercellular trafficking. This article is part of the theme issue 'Retrograde signalling from endosymbiotic organelles'.
Keywords: chloroplast; gun; intercellular trafficking; plasmodesmata; retrograde signalling; tetrapyrrole.
Conflict of interest statement
We declare we have no competing interests.
Figures





References
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
