

CD19-targeting fusion protein combined with PD1 antibody enhances anti-tumor immunity in mouse models
- PMID: 32363119
- PMCID: PMC7185221
- DOI: 10.1080/2162402X.2020.1747688
CD19-targeting fusion protein combined with PD1 antibody enhances anti-tumor immunity in mouse models
Abstract
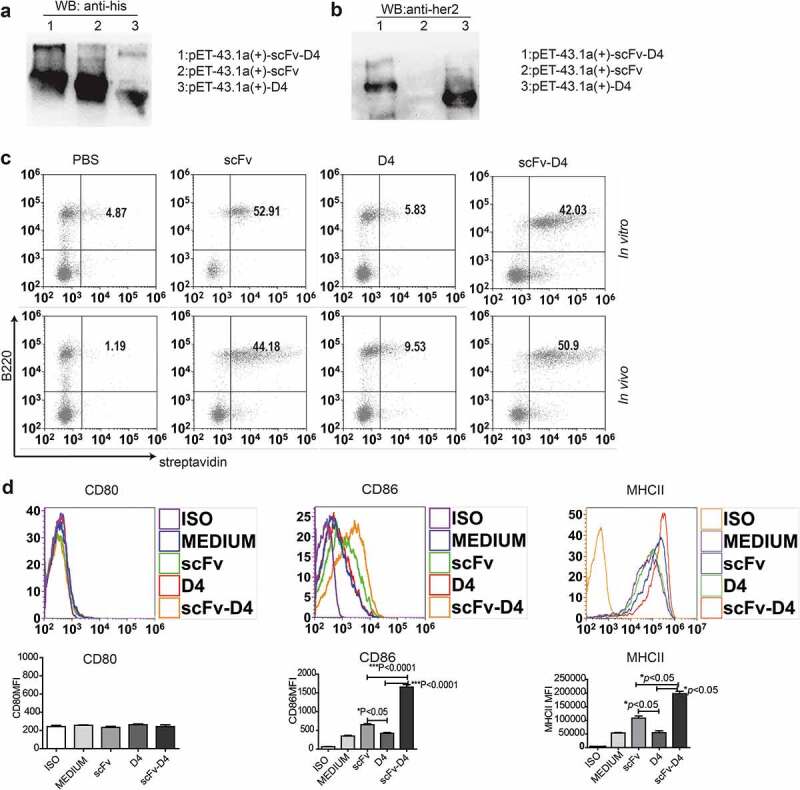
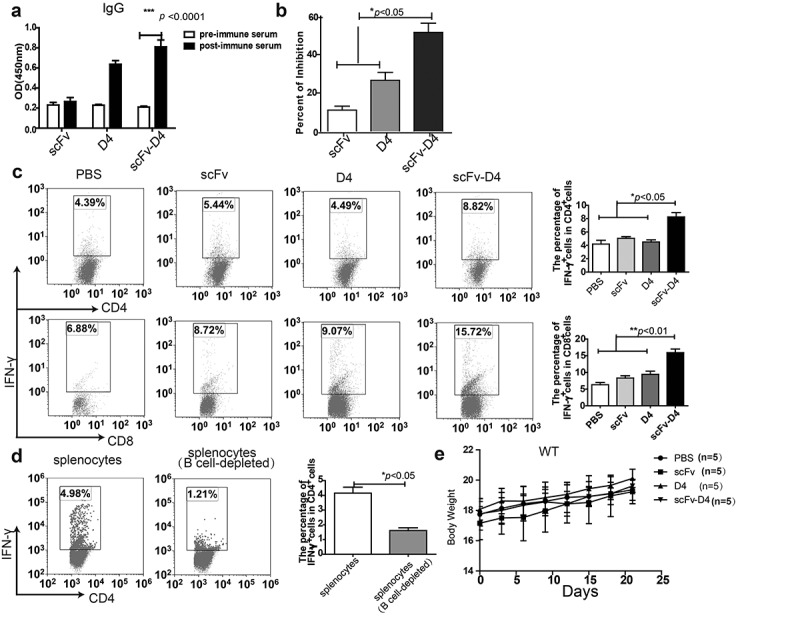
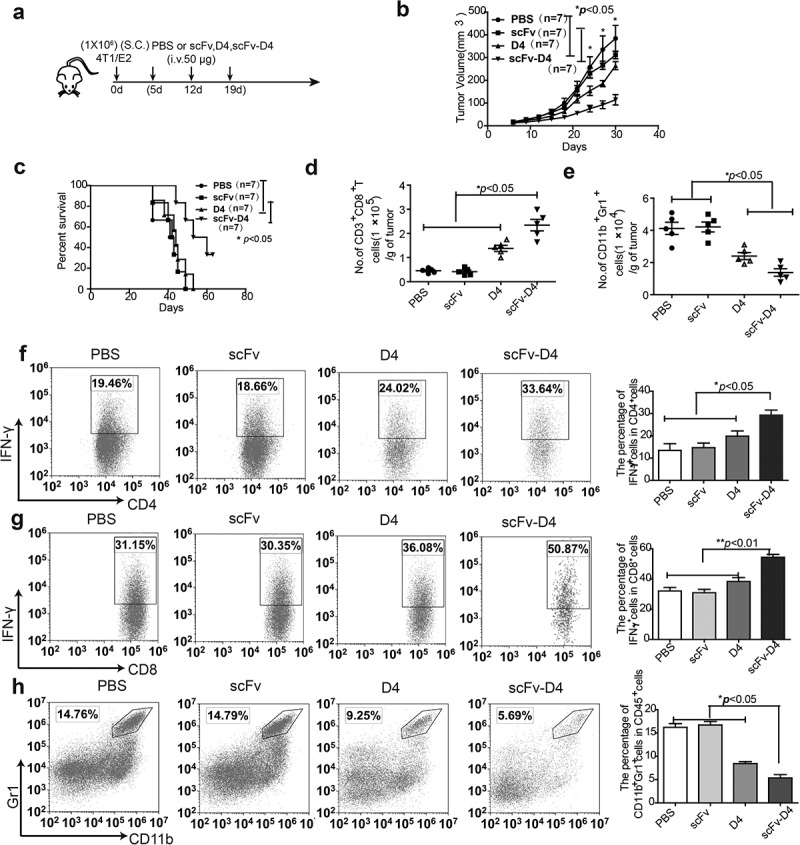
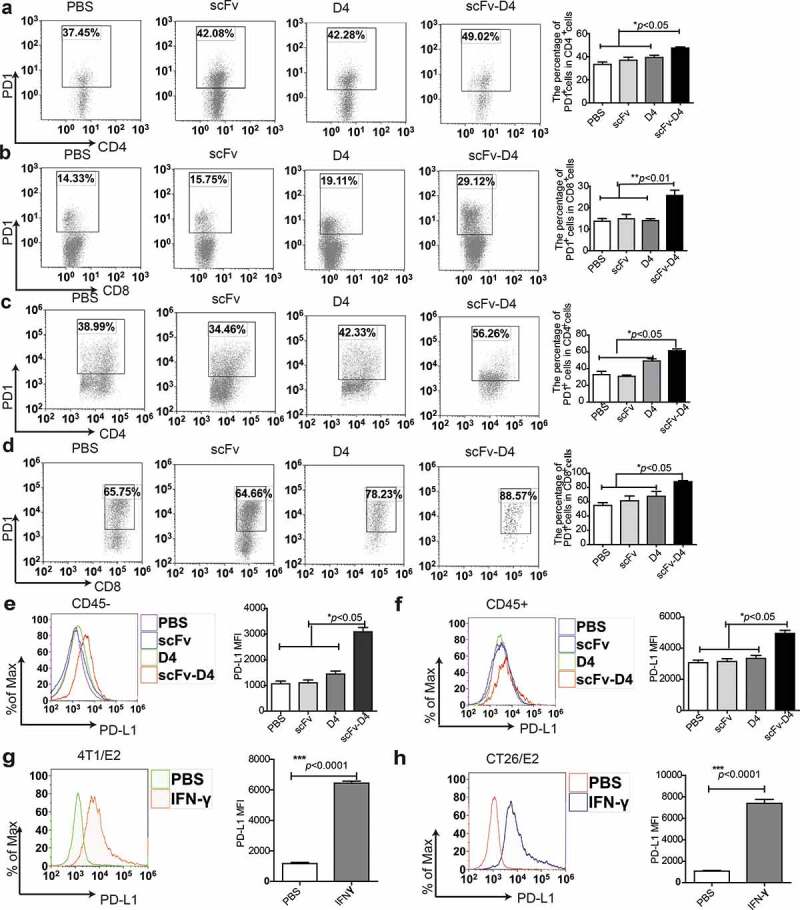
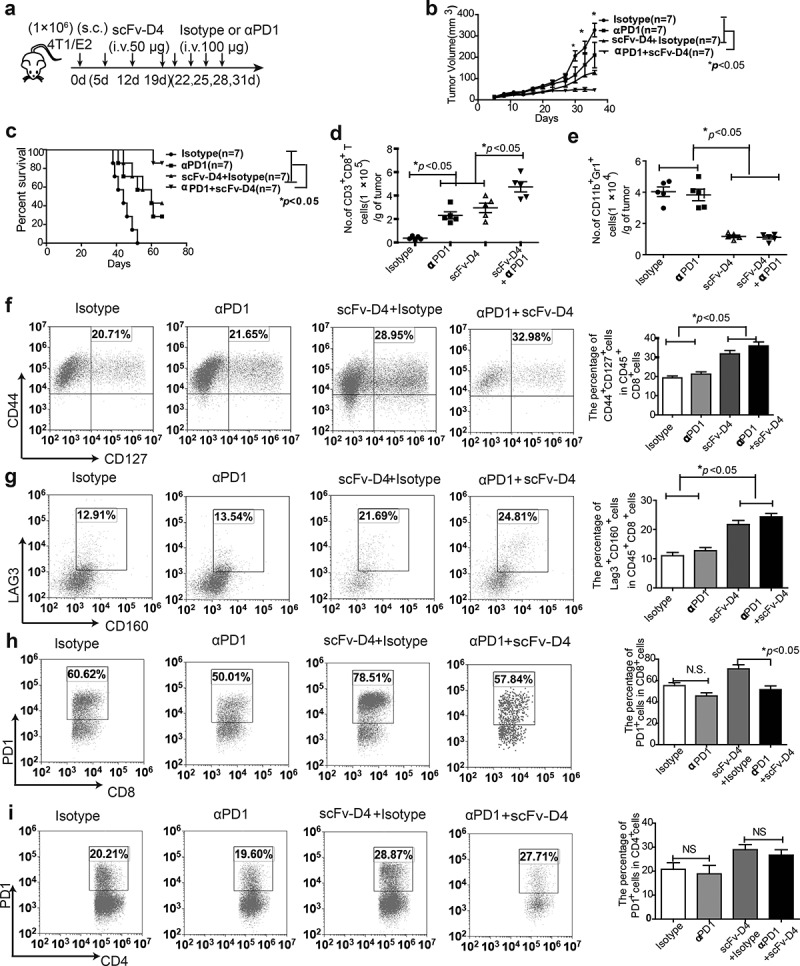
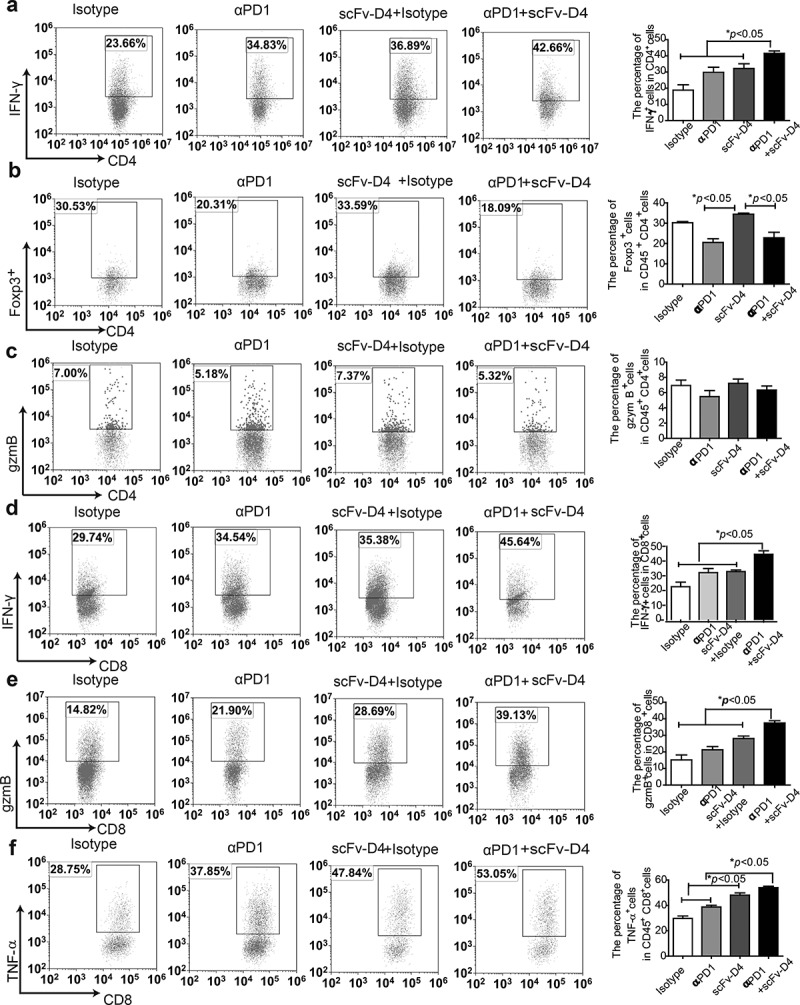
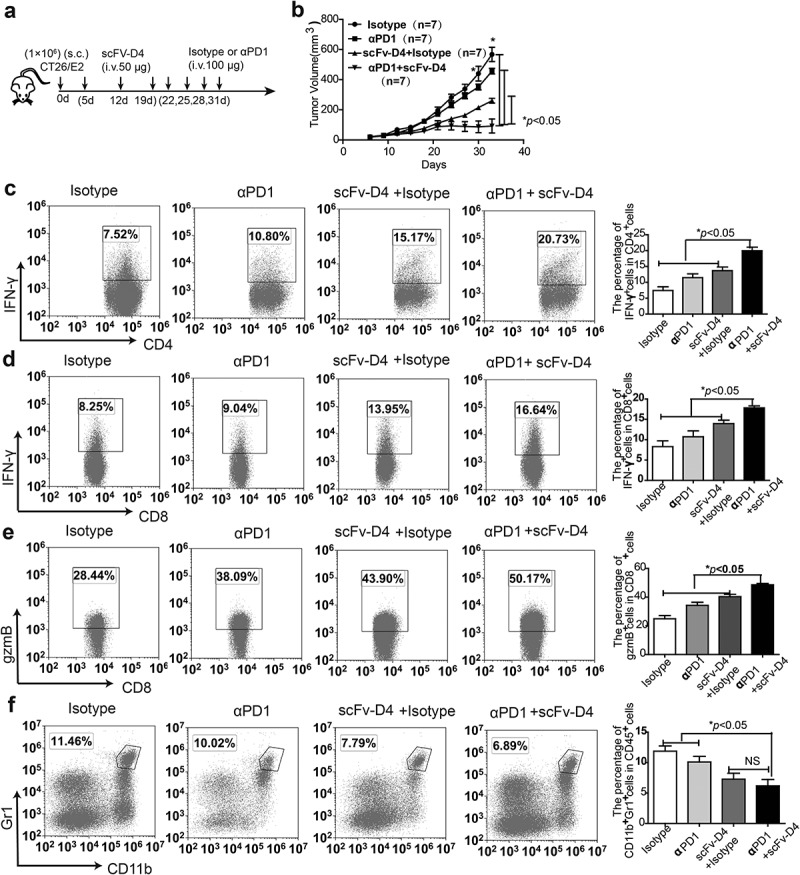
In our previous studies, using a B cell vaccine (scFv-Her2), the targeting of tumor-associated antigen Her2 (human epidermal growth factor receptor-2) to B cells via the anti-CD19 single chain variable fragment (scFv) was shown to augment tumor-specific immunity, which enhanced tumor control in the prophylactic and therapeutic setting. However, the fusion protein displayed limited activity against established tumors, and local relapses often occurred following scFv-Her2 treatment, indicating that scFv-Her2-induced responses are inadequate to maintain anti-tumor immunity. In this study, targeting the IV region (D4) of the extracellular region of Her2 to B cells via CD19 molecules (scFv-Her2D4) was found to enhance IFN-γ-producing-CD8+ T cell infiltration in tumor tissues and reduced the number of tumor-infiltrating myeloid-derived suppressor cells (MDSCs). However, negative co-stimulatory molecules such as programmed cell death protein-1 (PD-1), CD160, and LAG-3 on T cells and programmed death protein ligand-1 (PD-L1) on tumor cells were upregulated in the tumor microenvironment after scFv-Her2D4 treatment. Further, anti-PD1 administration enhanced the efficacy of scFv-Her2D4 and anti-tumor immunity, as evidenced by the reversal of tumor-infiltrating CD8+ T cell exhaustion and the reduction of MDSCs and Treg cells, which suppress T cells and alter the tumor immune microenvironment. Moreover, combining this with anti-PD1 antibodies promoted complete tumor rejection. Our data provide evidence of a close interaction among tumor vaccines, T cells, and the PD-L1/PD-1 axis and establish a basis for the rational design of combination therapy with immune modulators and tumor vaccine therapy.
Keywords: B cell vaccine; CD19; anti-PD1; breast cancer; combination therapy; immunotherapy.
© 2020 The Author(s). Published with license by Taylor & Francis Group, LLC.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous