

Loss of Snord116 impacts lateral hypothalamus, sleep, and food-related behaviors
- PMID: 32365348
- PMCID: PMC7406246
- DOI: 10.1172/jci.insight.137495
Loss of Snord116 impacts lateral hypothalamus, sleep, and food-related behaviors
Abstract
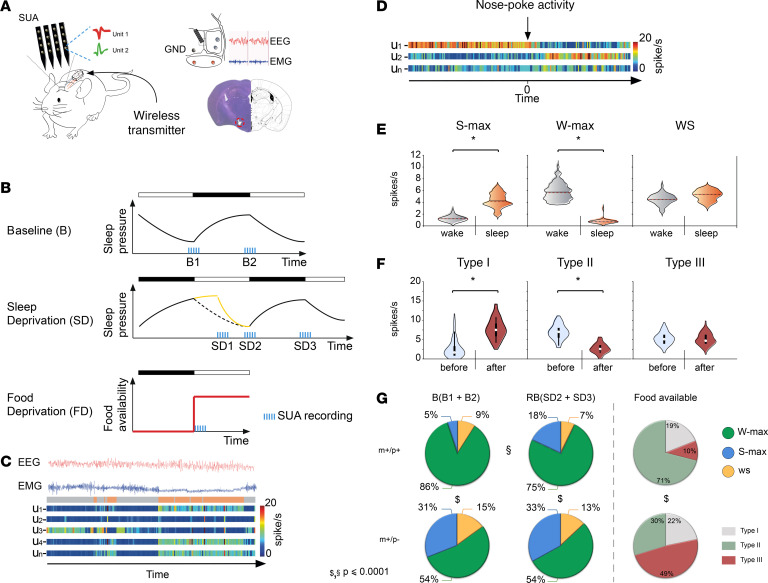
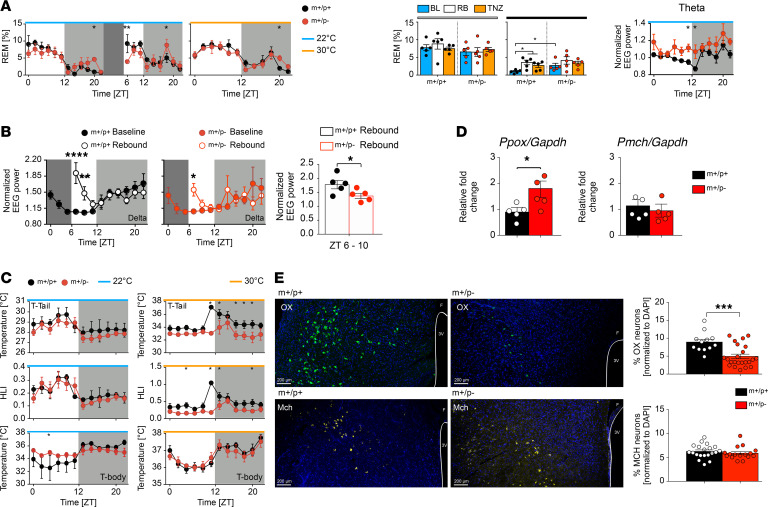
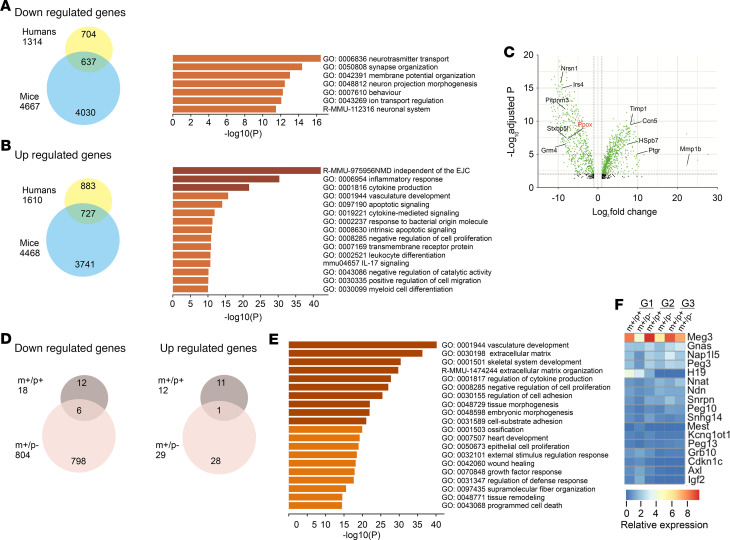
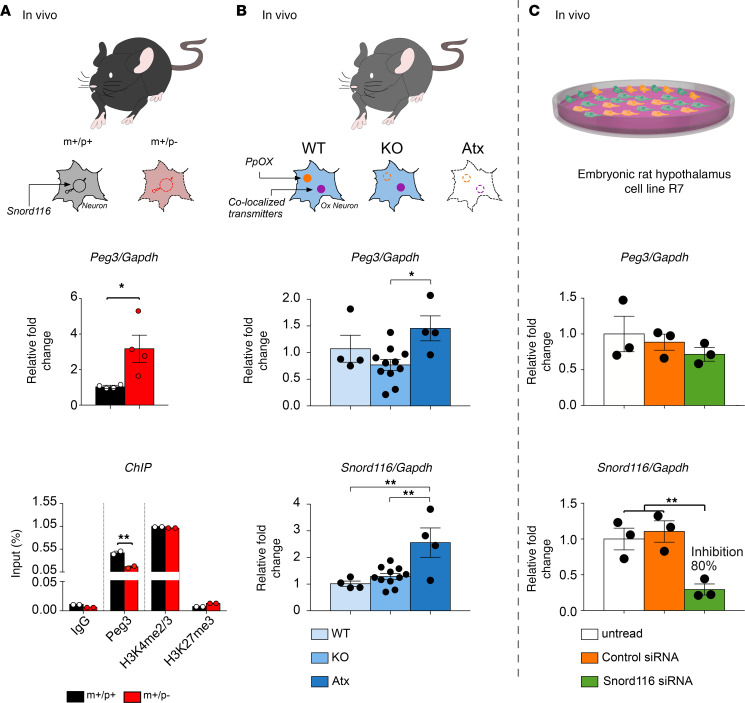
Imprinted genes are highly expressed in the hypothalamus; however, whether specific imprinted genes affect hypothalamic neuromodulators and their functions is unknown. It has been suggested that Prader-Willi syndrome (PWS), a neurodevelopmental disorder caused by lack of paternal expression at chromosome 15q11-q13, is characterized by hypothalamic insufficiency. Here, we investigate the role of the paternally expressed Snord116 gene within the context of sleep and metabolic abnormalities of PWS, and we report a significant role of this imprinted gene in the function and organization of the 2 main neuromodulatory systems of the lateral hypothalamus (LH) - namely, the orexin (OX) and melanin concentrating hormone (MCH) - systems. We observed that the dynamics between neuronal discharge in the LH and the sleep-wake states of mice with paternal deletion of Snord116 (PWScrm+/p-) are compromised. This abnormal state-dependent neuronal activity is paralleled by a significant reduction in OX neurons in the LH of mutant mice. Therefore, we propose that an imbalance between OX- and MCH-expressing neurons in the LH of mutant mice reflects a series of deficits manifested in the PWS, such as dysregulation of rapid eye movement (REM) sleep, food intake, and temperature control.
Keywords: Epigenetics; Neuroscience.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
