

Vascular Lipidomic Profiling of Potential Endogenous Fatty Acid PPAR Ligands Reveals the Coronary Artery as Major Producer of CYP450-Derived Epoxy Fatty Acids
- PMID: 32365470
- PMCID: PMC7290345
- DOI: 10.3390/cells9051096
Vascular Lipidomic Profiling of Potential Endogenous Fatty Acid PPAR Ligands Reveals the Coronary Artery as Major Producer of CYP450-Derived Epoxy Fatty Acids
Abstract
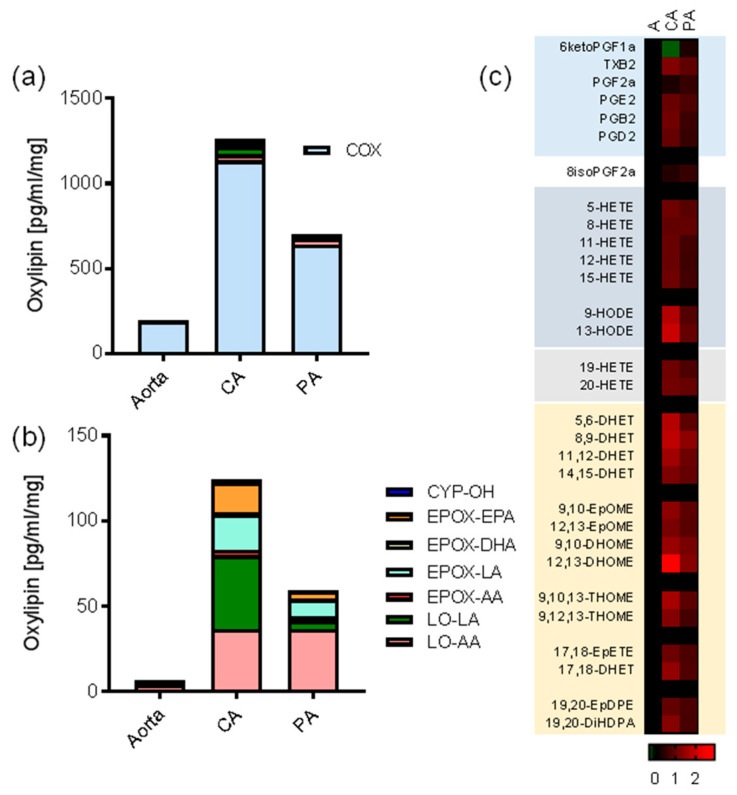
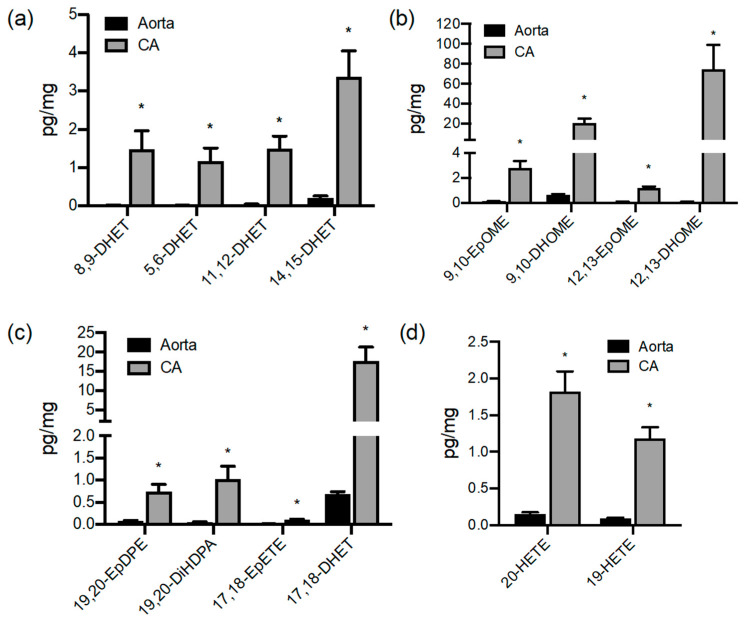
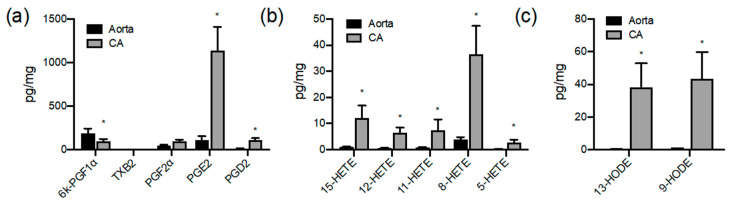
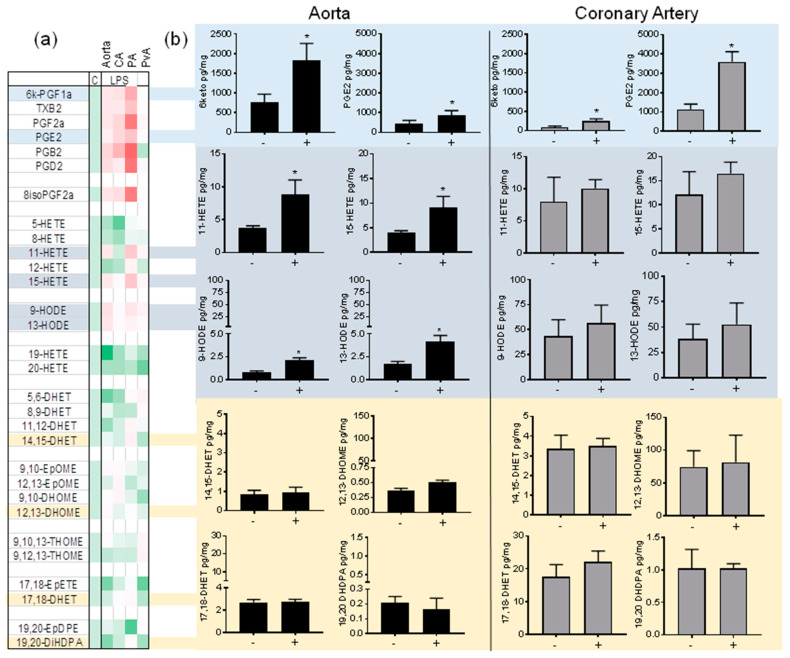
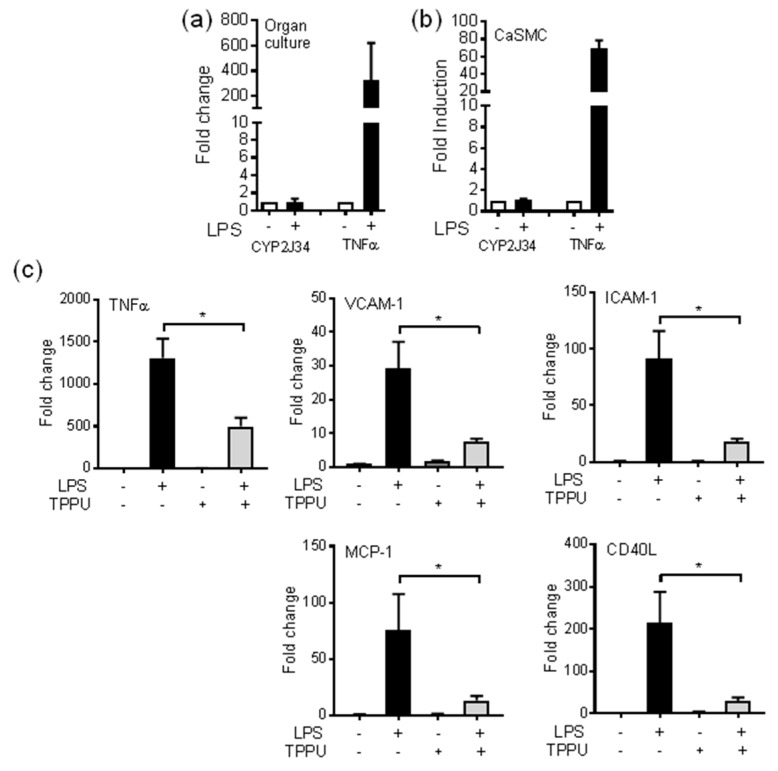
A number of oxylipins have been described as endogenous PPAR ligands. The very short biological half-lives of oxylipins suggest roles as autocrine or paracrine signaling molecules. While coronary arterial atherosclerosis is the root of myocardial infarction, aortic atherosclerotic plaque formation is a common readout of in vivo atherosclerosis studies in mice. Improved understanding of the compartmentalized sources of oxylipin PPAR ligands will increase our knowledge of the roles of PPAR signaling in diverse vascular tissues. Here, we performed a targeted lipidomic analysis of ex vivo-generated oxylipins from porcine aorta, coronary artery, pulmonary artery and perivascular adipose. Cyclooxygenase (COX)-derived prostanoids were the most abundant detectable oxylipin from all tissues. By contrast, the coronary artery produced significantly higher levels of oxylipins from CYP450 pathways than other tissues. The TLR4 ligand LPS induced prostanoid formation in all vascular tissue tested. The 11-HETE, 15-HETE, and 9-HODE were also induced by LPS from the aorta and pulmonary artery but not coronary artery. Epoxy fatty acid (EpFA) formation was largely unaffected by LPS. The pig CYP2J homologue CYP2J34 was expressed in porcine vascular tissue and primary coronary artery smooth muscle cells (pCASMCs) in culture. Treatment of pCASMCs with LPS induced a robust profile of pro-inflammatory target genes: TNFα, ICAM-1, VCAM-1, MCP-1 and CD40L. The soluble epoxide hydrolase inhibitor TPPU, which prevents the breakdown of endogenous CYP-derived EpFAs, significantly suppressed LPS-induced inflammatory target genes. In conclusion, PPAR-activating oxylipins are produced and regulated in a vascular site-specific manner. The CYP450 pathway is highly active in the coronary artery and capable of providing anti-inflammatory oxylipins that prevent processes of inflammatory vascular disease progression.
Keywords: CYP450; PPARs; coronary artery; eicosanoids; inflammation; lipidomics; vascular.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
References
-
- Bailey J.M., Bryant R.W., Whiting J., Salata K. Characterization of 11-HETE and 15-HETE, together with prostacyclin, as major products of the cyclooxygenase pathway in cultured rat aorta smooth muscle cells. J. Lipid Res. 1983;24:1419–1428. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
