

Turbulent coherent structures and early life below the Kolmogorov scale
- PMID: 32366844
- PMCID: PMC7198613
- DOI: 10.1038/s41467-020-15780-1
Turbulent coherent structures and early life below the Kolmogorov scale
Abstract
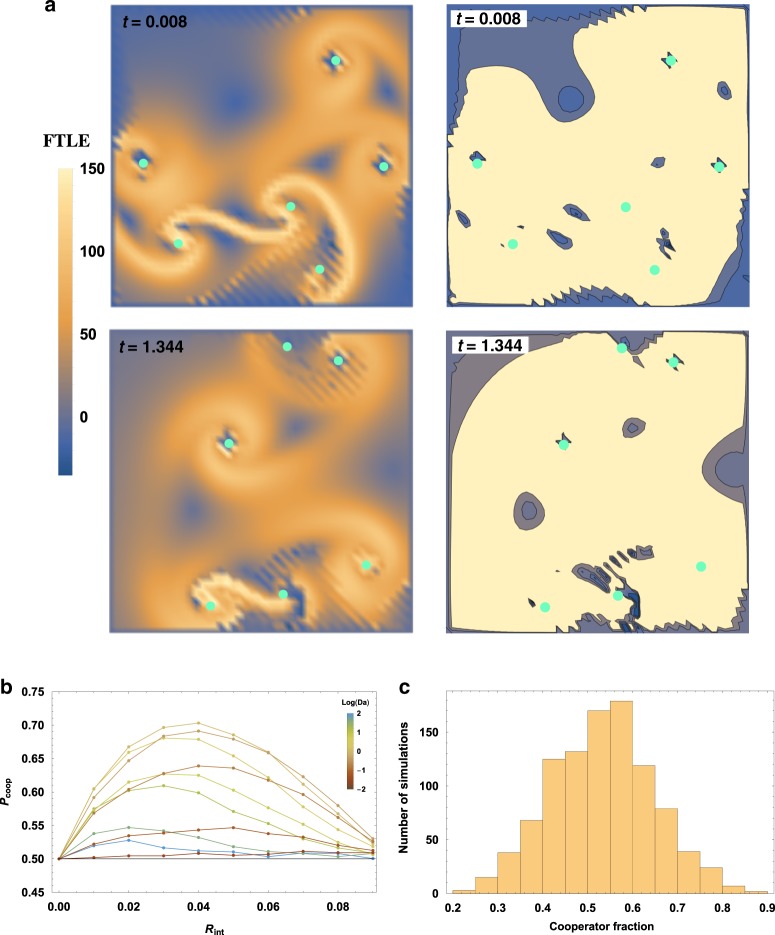
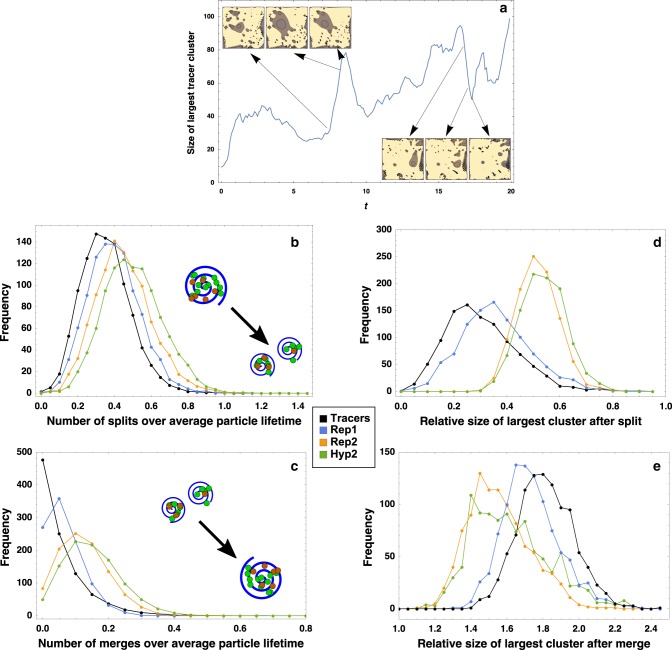
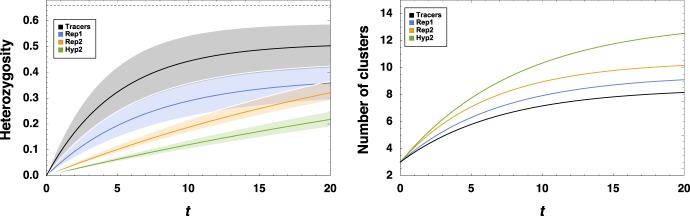
Major evolutionary transitions, including the emergence of life, likely occurred in aqueous environments. While the role of water's chemistry in early life is well studied, the effects of water's ability to manipulate population structure are less clear. Population structure is known to be critical, as effective replicators must be insulated from parasites. Here, we propose that turbulent coherent structures, long-lasting flow patterns which trap particles, may serve many of the properties associated with compartments - collocalization, division, and merging - which are commonly thought to play a key role in the origins of life and other evolutionary transitions. We substantiate this idea by simulating multiple proposed metabolisms for early life in a simple model of a turbulent flow, and find that balancing the turnover times of biological particles and coherent structures can indeed enhance the likelihood of these metabolisms overcoming extinction either via parasitism or via a lack of metabolic support. Our results suggest that group selection models may be applicable with fewer physical and chemical constraints than previously thought, and apply much more widely in aqueous environments.
Conflict of interest statement
The authors declare no competing interests.
Figures
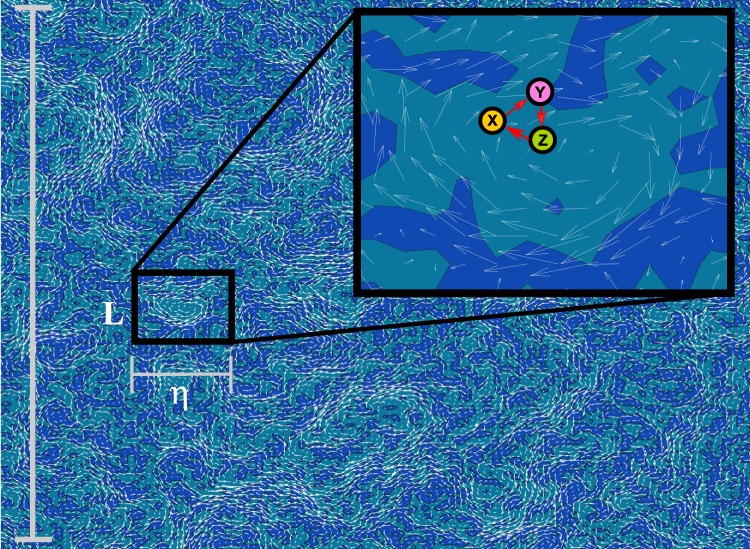
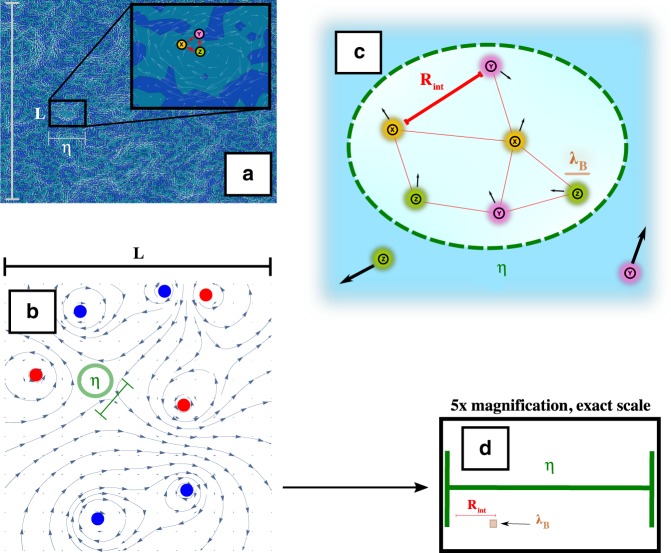
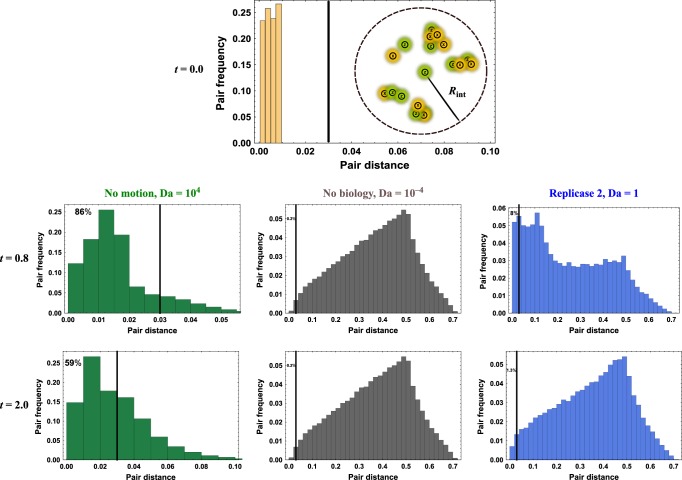
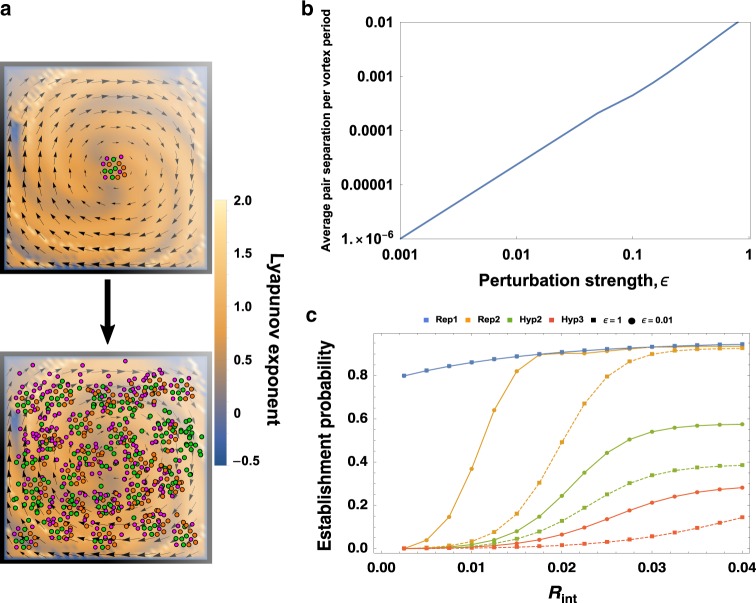
References
-
- Smith, J. M. & Szathmary, E. The Major Transitions in Evolution (Oxford University Press, 1997).
-
- Zwicker D, Seyboldt R, Weber CA, Hyman AA, Jülicher F. Growth and division of active droplets provides a model for protocells. Nat. Phys. 2017;13:408. doi: 10.1038/nphys3984. - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
