

Multimodal determinants of phase-locked dynamics across deep-superficial hippocampal sublayers during theta oscillations
- PMID: 32371879
- PMCID: PMC7200700
- DOI: 10.1038/s41467-020-15840-6
Multimodal determinants of phase-locked dynamics across deep-superficial hippocampal sublayers during theta oscillations
Abstract
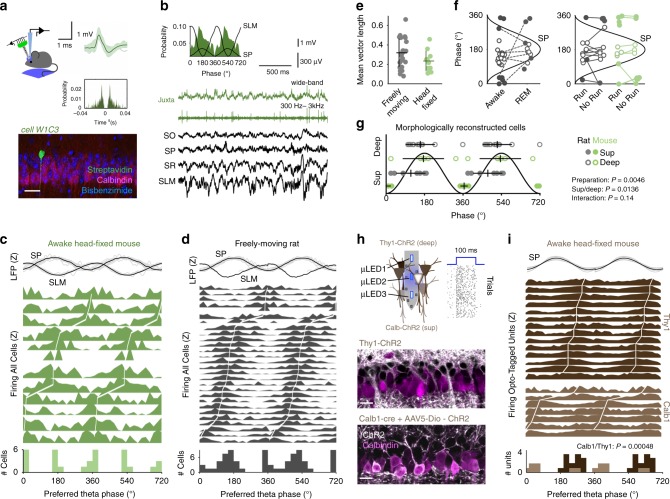
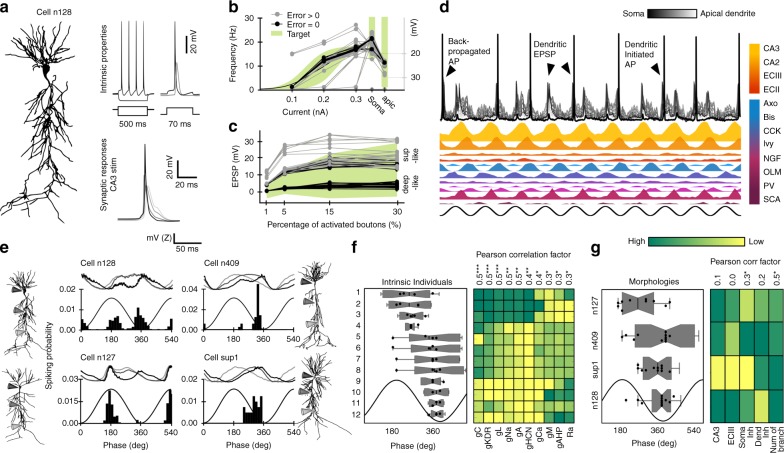
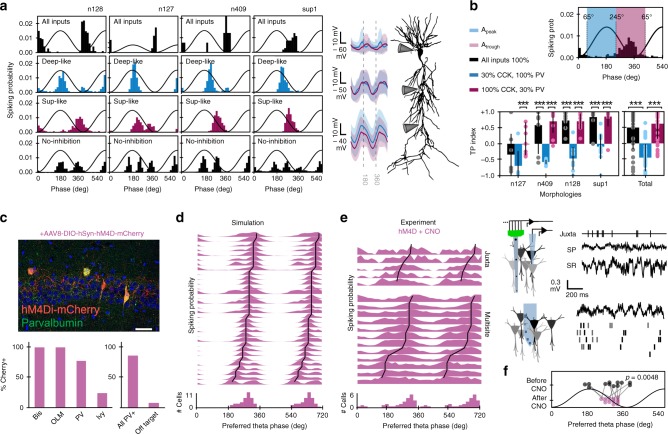
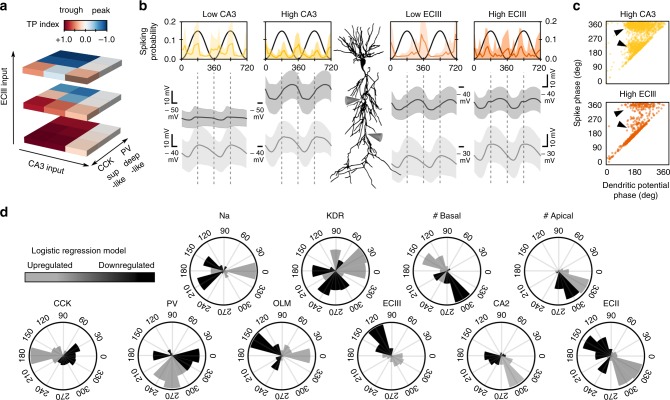
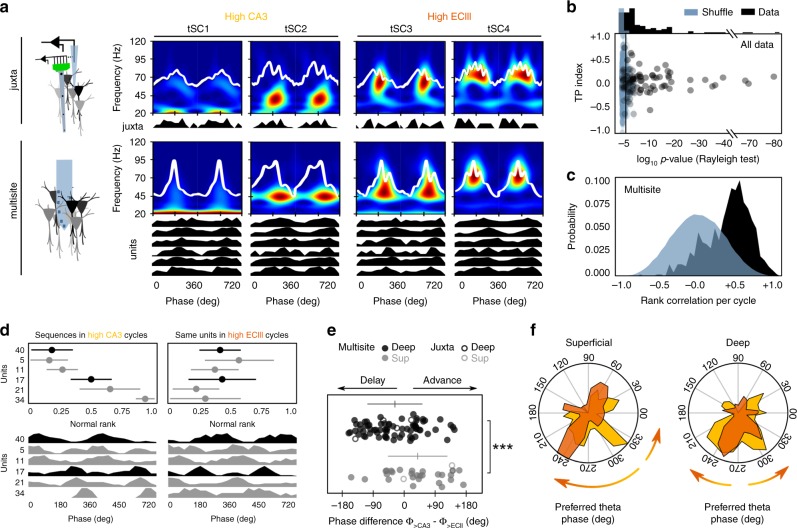
Theta oscillations play a major role in temporarily defining the hippocampal rate code by translating behavioral sequences into neuronal representations. However, mechanisms constraining phase timing and cell-type-specific phase preference are unknown. Here, we employ computational models tuned with evolutionary algorithms to evaluate phase preference of individual CA1 pyramidal cells recorded in mice and rats not engaged in any particular memory task. We applied unbiased and hypothesis-free approaches to identify effects of intrinsic and synaptic factors, as well as cell morphology, in determining phase preference. We found that perisomatic inhibition delivered by complementary populations of basket cells interacts with input pathways to shape phase-locked specificity of deep and superficial pyramidal cells. Somatodendritic integration of fluctuating glutamatergic inputs defined cycle-by-cycle by unsupervised methods demonstrated that firing selection is tuneable across sublayers. Our data identify different mechanisms of phase-locking selectivity that are instrumental for flexible dynamical representations of theta sequences.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Excitatory Inputs Determine Phase-Locking Strength and Spike-Timing of CA1 Stratum Oriens/Alveus Parvalbumin and Somatostatin Interneurons during Intrinsically Generated Hippocampal Theta Rhythm.J Neurosci. 2016 Jun 22;36(25):6605-22. doi: 10.1523/JNEUROSCI.3951-13.2016. J Neurosci. 2016. PMID: 27335395 Free PMC article.
-
A computational study on plasticity during theta cycles at Schaffer collateral synapses on CA1 pyramidal cells in the hippocampus.Hippocampus. 2015 Feb;25(2):208-18. doi: 10.1002/hipo.22365. Epub 2014 Sep 25. Hippocampus. 2015. PMID: 25220633
-
Intrinsic Mechanisms of Frequency Selectivity in the Proximal Dendrites of CA1 Pyramidal Neurons.J Neurosci. 2018 Sep 19;38(38):8110-8127. doi: 10.1523/JNEUROSCI.0449-18.2018. Epub 2018 Aug 3. J Neurosci. 2018. PMID: 30076213 Free PMC article.
-
Associative memory storage and retrieval: involvement of theta oscillations in hippocampal information processing.Neural Plast. 2011;2011:683961. doi: 10.1155/2011/683961. Epub 2011 Sep 26. Neural Plast. 2011. PMID: 21961072 Free PMC article. Review.
-
Allocentric representations of space in the hippocampus.Neurosci Res. 2020 Apr;153:1-7. doi: 10.1016/j.neures.2019.06.002. Epub 2019 Jul 2. Neurosci Res. 2020. PMID: 31276699 Review.
Cited by
-
Silencing KCC2 in mouse dorsal hippocampus compromises spatial and contextual memory.Neuropsychopharmacology. 2023 Jun;48(7):1067-1077. doi: 10.1038/s41386-022-01480-5. Epub 2022 Oct 27. Neuropsychopharmacology. 2023. PMID: 36302847 Free PMC article.
-
Potential factors influencing replay across CA1 during sharp-wave ripples.Philos Trans R Soc Lond B Biol Sci. 2020 May 25;375(1799):20190236. doi: 10.1098/rstb.2019.0236. Epub 2020 Apr 6. Philos Trans R Soc Lond B Biol Sci. 2020. PMID: 32248778 Free PMC article.
-
Spiking Neural Networks and Hippocampal Function: A Web-Accessible Survey of Simulations, Modeling Methods, and Underlying Theories.Cogn Syst Res. 2021 Dec;70:80-92. doi: 10.1016/j.cogsys.2021.07.008. Epub 2021 Jul 31. Cogn Syst Res. 2021. PMID: 34504394 Free PMC article.
-
Integrating new memories into the hippocampal network activity space.Nat Neurosci. 2021 Mar;24(3):326-330. doi: 10.1038/s41593-021-00804-w. Epub 2021 Feb 18. Nat Neurosci. 2021. PMID: 33603228 Free PMC article.
-
Months-long tracking of neuronal ensembles spanning multiple brain areas with Ultra-Flexible Tentacle Electrodes.Nat Commun. 2024 Jun 6;15(1):4822. doi: 10.1038/s41467-024-49226-9. Nat Commun. 2024. PMID: 38844769 Free PMC article.
References
-
- Arieli A, Sterkin A, Grinvald A, Aertsen A. Dynamics of ongoing activity: explanation of the large variability in evoked cortical responses. Science. 1996;273:1868–1871. - PubMed
-
- Dragoi G, Buzsáki G. Temporal encoding of place sequences by hippocampal cell assemblies. Neuron. 2006;50:145–157. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
