

The F220C and F45L rhodopsin mutations identified in retinitis pigmentosa patients do not cause pathology in mice
- PMID: 32371886
- PMCID: PMC7200662
- DOI: 10.1038/s41598-020-64437-y
The F220C and F45L rhodopsin mutations identified in retinitis pigmentosa patients do not cause pathology in mice
Abstract
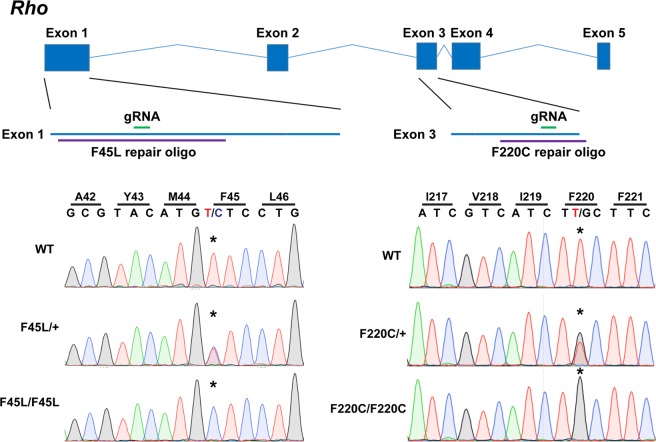
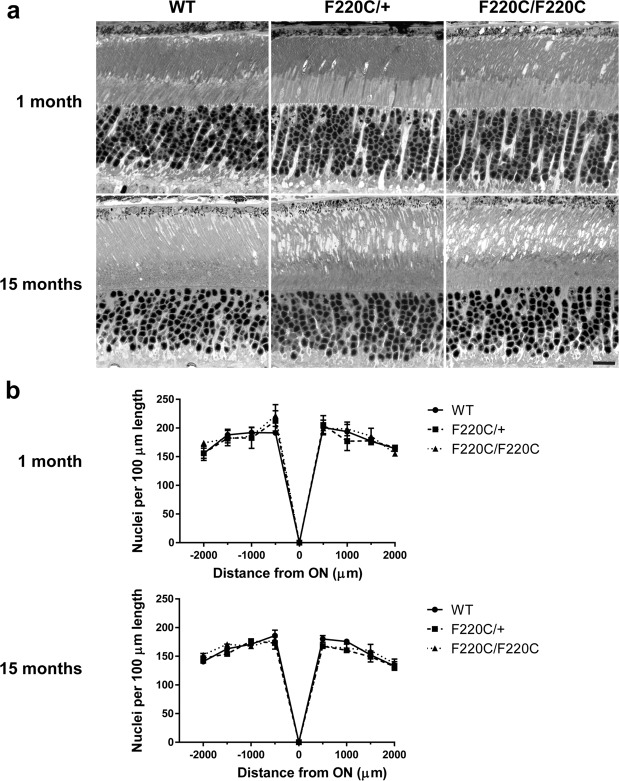
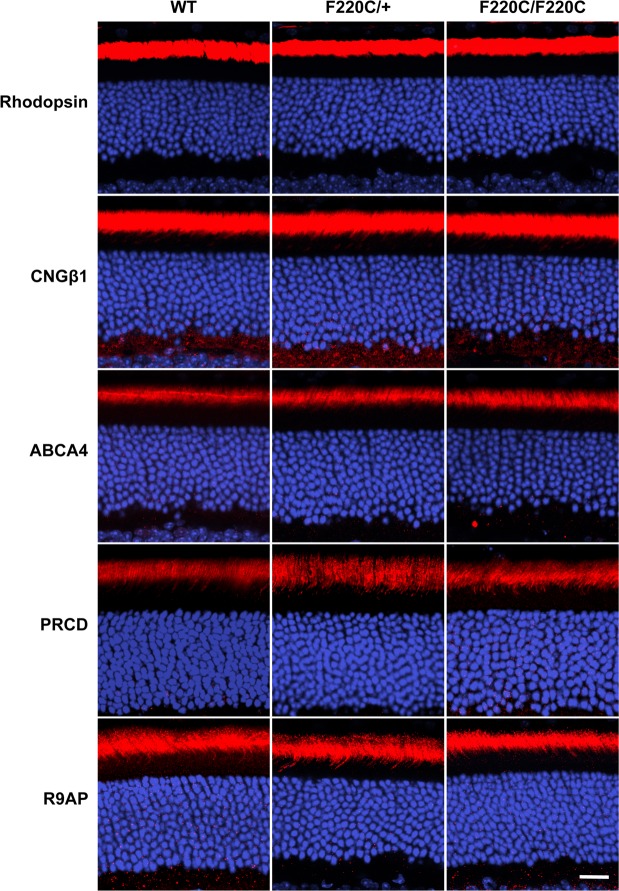
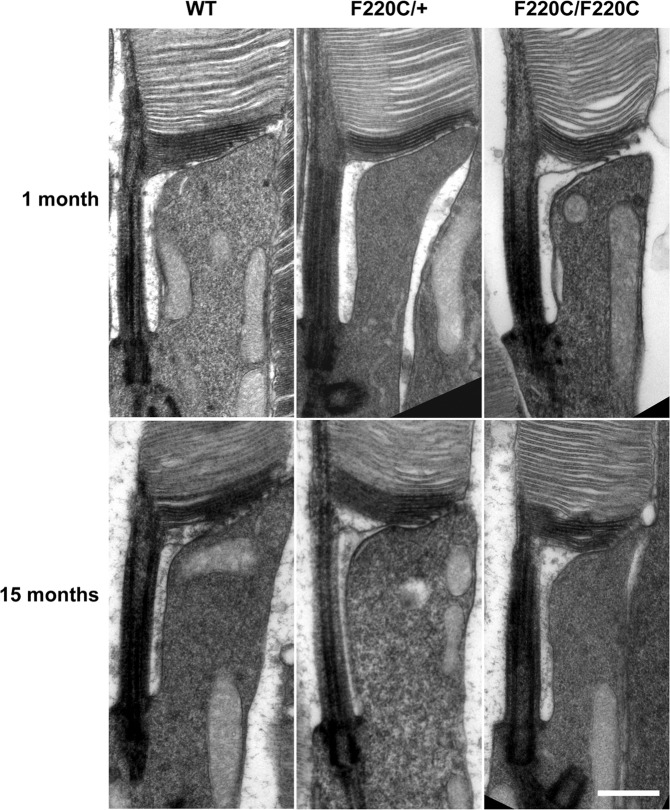
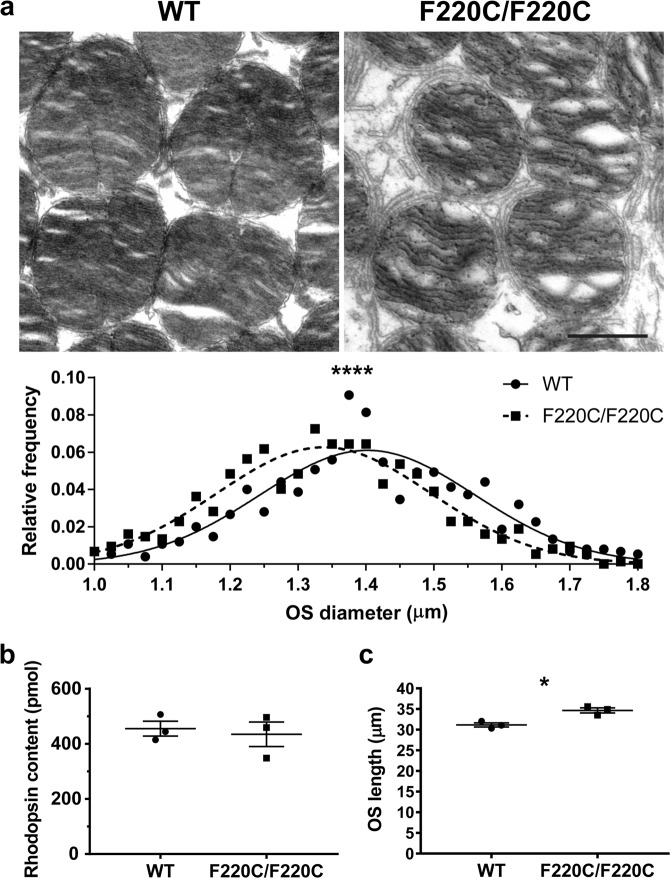
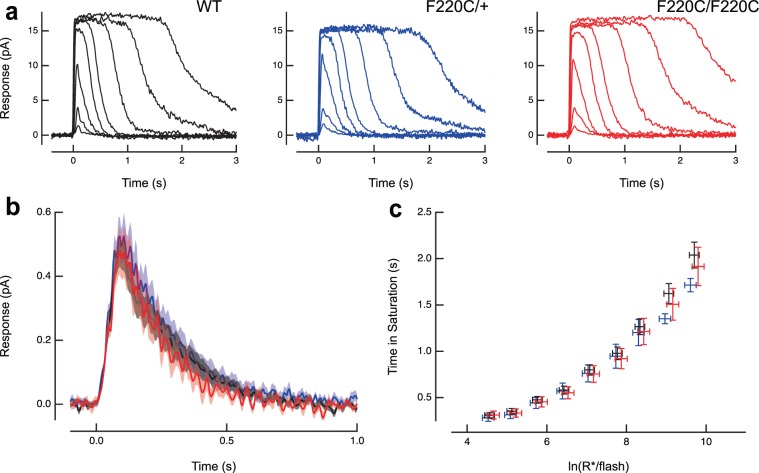
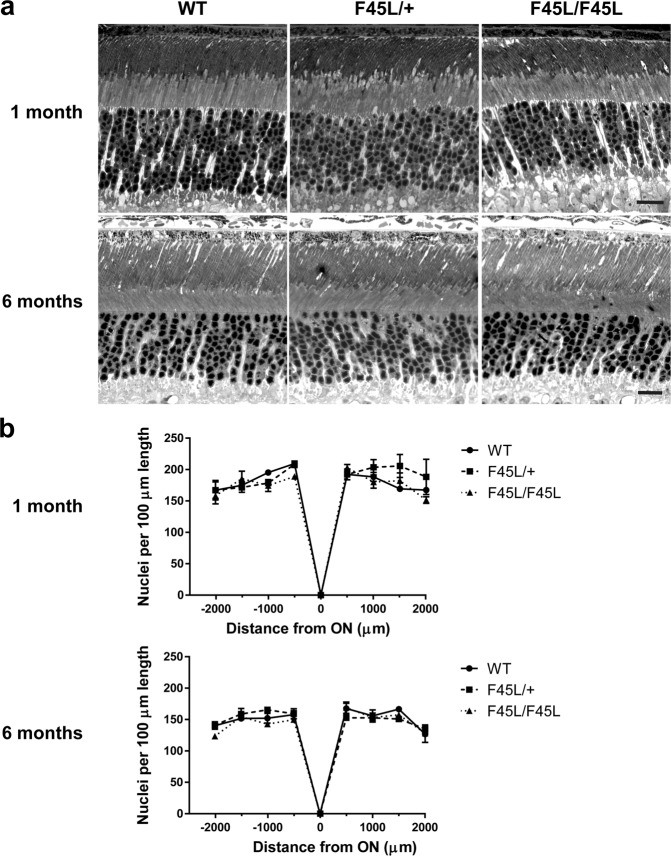
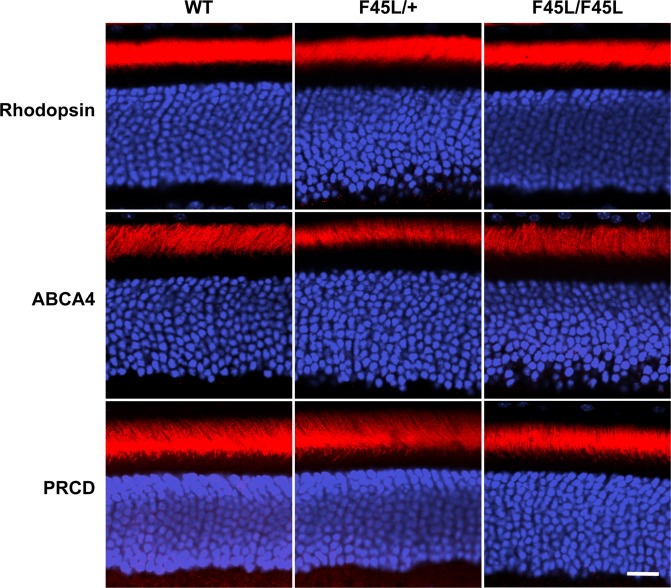
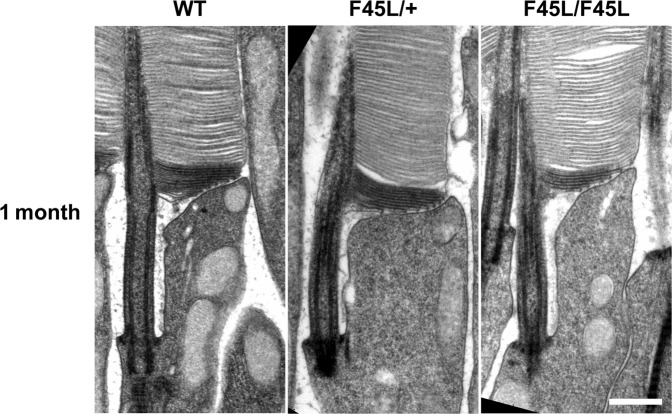
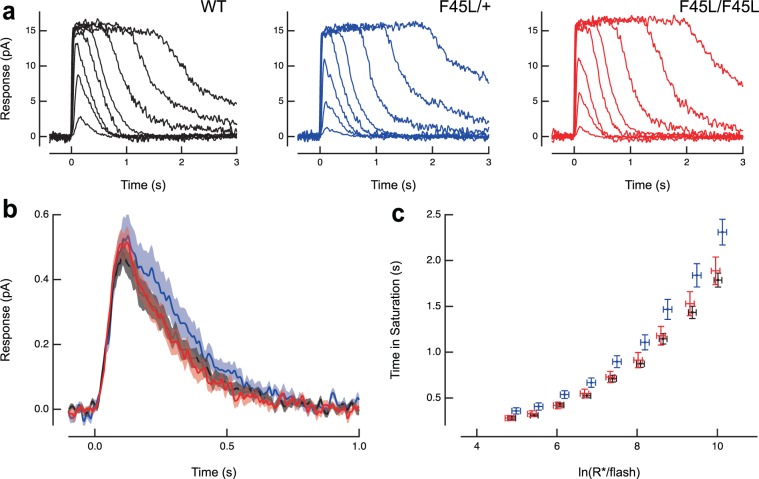
Retinitis pigmentosa is a retinal degenerative disease that leads to blindness through photoreceptor loss. Rhodopsin is the most frequently mutated protein in this disease. While many rhodopsin mutations have well-understood consequences that lead to cell death, the disease association of several rhodopsin mutations identified in retinitis pigmentosa patients, including F220C and F45L, has been disputed. In this study, we generated two knockin mouse lines bearing each of these mutations. We did not observe any photoreceptor degeneration in either heterozygous or homozygous animals of either line. F220C mice exhibited minor disruptions of photoreceptor outer segment dimensions without any mislocalization of outer segment proteins, whereas photoreceptors of F45L mice were normal. Suction electrode recordings from individual photoreceptors of both mutant lines showed normal flash sensitivity and photoresponse kinetics. Taken together, these data suggest that neither the F220C nor F45L mutation has pathological consequences in mice and, therefore, may not be causative of retinitis pigmentosa in humans.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Dimerization deficiency of enigmatic retinitis pigmentosa-linked rhodopsin mutants.Nat Commun. 2016 Oct 3;7:12832. doi: 10.1038/ncomms12832. Nat Commun. 2016. PMID: 27694816 Free PMC article.
-
The severe autosomal dominant retinitis pigmentosa rhodopsin mutant Ter349Glu mislocalizes and induces rapid rod cell death.J Biol Chem. 2013 Oct 4;288(40):29047-55. doi: 10.1074/jbc.M113.495184. Epub 2013 Aug 12. J Biol Chem. 2013. PMID: 23940033 Free PMC article.
-
A diffusible factor from normal retinal cells promotes rod photoreceptor survival in an in vitro model of retinitis pigmentosa.J Neurobiol. 1999 Jun 15;39(4):475-90. J Neurobiol. 1999. PMID: 10380070
-
Defective trafficking of rhodopsin and its role in retinal degenerations.Int Rev Cell Mol Biol. 2012;293:1-44. doi: 10.1016/B978-0-12-394304-0.00006-3. Int Rev Cell Mol Biol. 2012. PMID: 22251557 Review.
-
Mechanisms of cell death in rhodopsin retinitis pigmentosa: implications for therapy.Trends Mol Med. 2005 Apr;11(4):177-85. doi: 10.1016/j.molmed.2005.02.007. Trends Mol Med. 2005. PMID: 15823756 Review.
Cited by
-
Investigating the Role of Rhodopsin F45L Mutation in Mouse Rod Photoreceptor Signaling and Survival.eNeuro. 2023 Mar 7;10(3):ENEURO.0330-22.2023. doi: 10.1523/ENEURO.0330-22.2023. Print 2023 Mar. eNeuro. 2023. PMID: 36823167 Free PMC article.
-
Unraveling the genetic complexities of combined retinal dystrophy and hearing impairment.Hum Genet. 2022 Apr;141(3-4):785-803. doi: 10.1007/s00439-021-02303-1. Epub 2021 Jun 20. Hum Genet. 2022. PMID: 34148116 Free PMC article.
-
Supramolecular organization of rhodopsin in rod photoreceptor cell membranes.Pflugers Arch. 2021 Sep;473(9):1361-1376. doi: 10.1007/s00424-021-02522-5. Epub 2021 Feb 16. Pflugers Arch. 2021. PMID: 33591421 Free PMC article. Review.
-
Unusual mode of dimerization of retinitis pigmentosa-associated F220C rhodopsin.Sci Rep. 2021 May 18;11(1):10536. doi: 10.1038/s41598-021-90039-3. Sci Rep. 2021. PMID: 34006992 Free PMC article.
-
Photoreceptor disc incisures form as an adaptive mechanism ensuring the completion of disc enclosure.bioRxiv [Preprint]. 2023 Apr 7:2023.04.06.535932. doi: 10.1101/2023.04.06.535932. bioRxiv. 2023. Update in: Elife. 2023 Jul 14;12:e89160. doi: 10.7554/eLife.89160. PMID: 37066355 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
