

Human Cerebrospinal Fluid Induces Neuronal Excitability Changes in Resected Human Neocortical and Hippocampal Brain Slices
- PMID: 32372899
- PMCID: PMC7186381
- DOI: 10.3389/fnins.2020.00283
Human Cerebrospinal Fluid Induces Neuronal Excitability Changes in Resected Human Neocortical and Hippocampal Brain Slices
Abstract
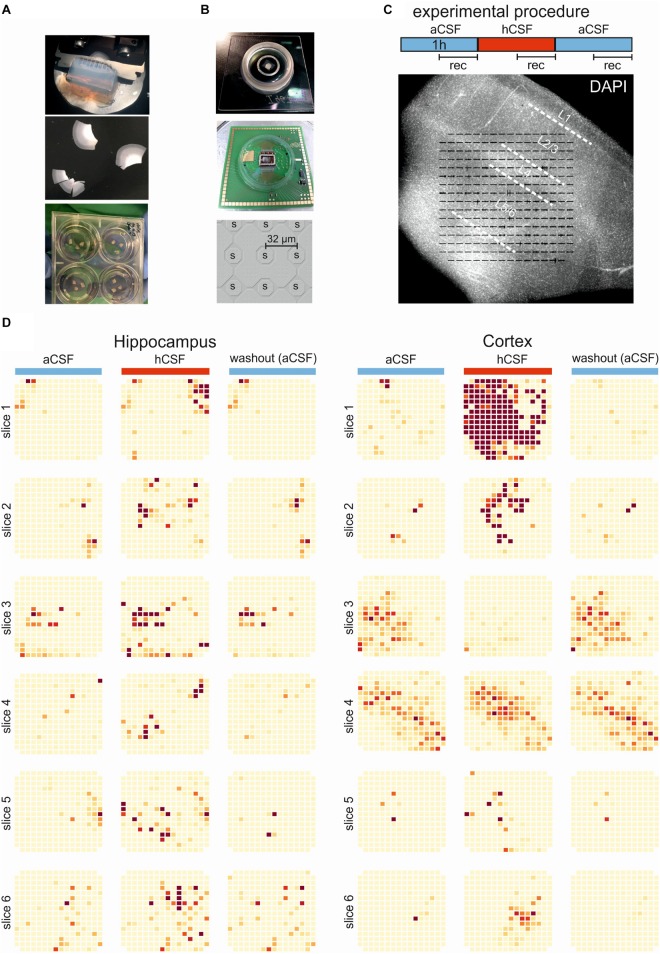
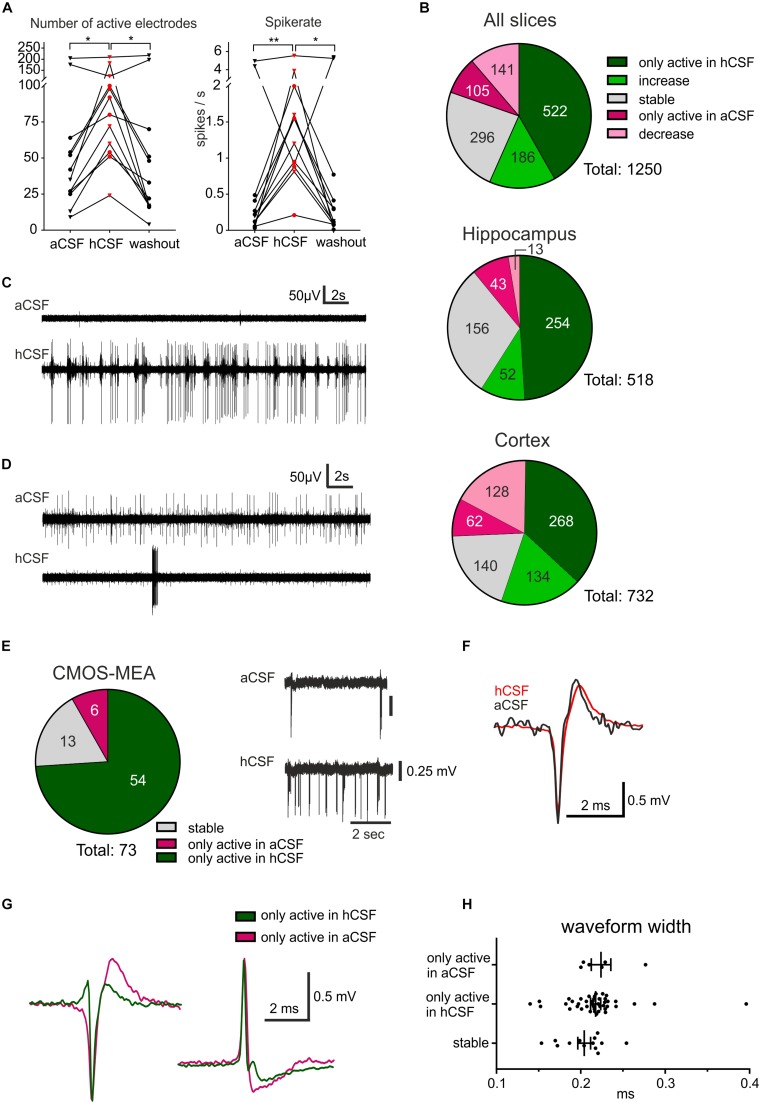
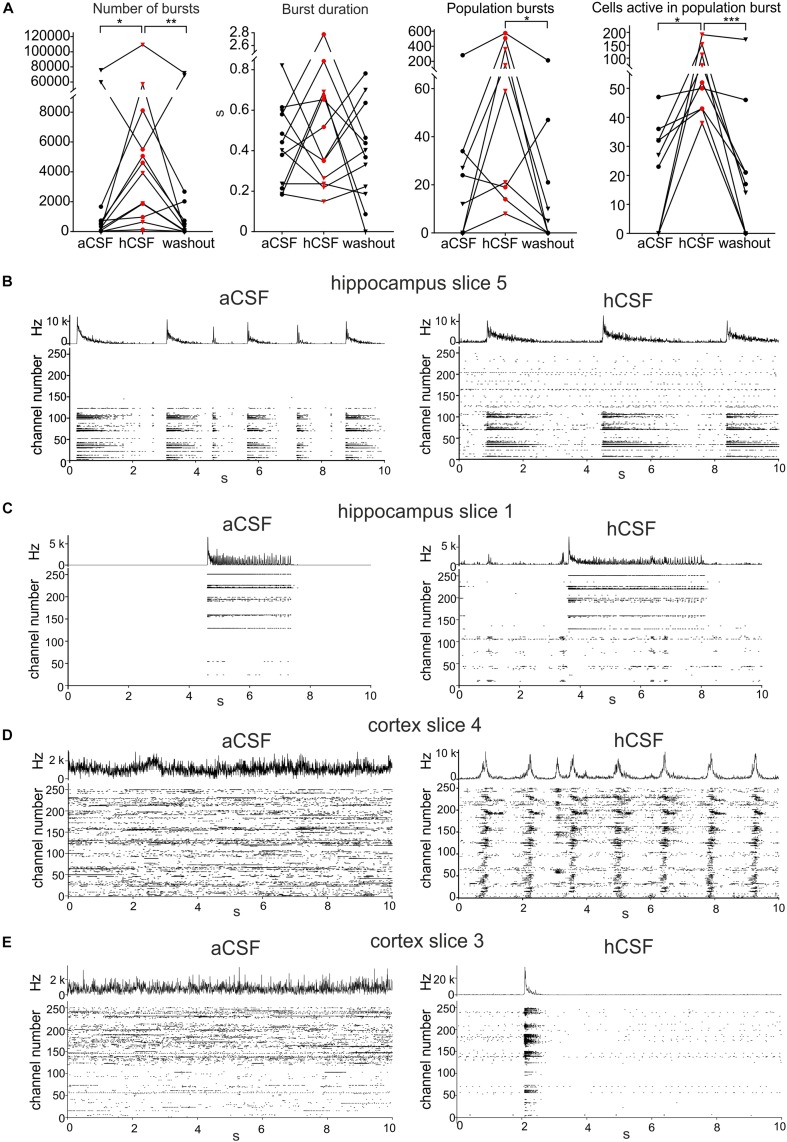
Human cerebrospinal fluid (hCSF) has proven advantageous over conventional medium for culturing both rodent and human brain tissue. In addition, increased activity and synchrony, closer to the dynamic states exclusively recorded in vivo, were reported in rodent slices and cell cultures switching from artificial cerebrospinal fluid (aCSF) to hCSF. This indicates that hCSF possesses properties that are not matched by the aCSF, which is generally used for most electrophysiological recordings. To evaluate the possible significance of using hCSF as an electrophysiological recording medium, also for human brain tissue, we compared the network and single-cell firing properties of human brain slice cultures during perfusion with hCSF and aCSF. For measuring the overall activity from a majority of neurons within neocortical and hippocampal human slices, we used a microelectrode array (MEA) recording technique with 252 electrodes covering an area of 3.2 × 3.2 mm2. A second CMOS-based MEA with 4225 sensors on a 2 × 2 mm2 area was used for detailed mapping of action potential waveforms and cell identification. We found that hCSF increased the number of active electrodes and neurons and the firing rate of the neurons in the slices and induced an increase in the numbers of single channel and population bursts. Interestingly, not only an increase in the overall activity in the slices was observed, but a reconfiguration of the network could also be detected with specific activation and inactivation of subpopulations of neuronal ensembles. In conclusion, hCSF is an important component to consider for future human brain slice studies, especially for experiments designed to mimic parts of physiology and disease observed in vivo.
Keywords: CMOS-MEA; cortex; hippocampus; human cerebrospinal fluid; human tissue; microelectrode array; organotypic slices.
Copyright © 2020 Wickham, Corna, Schwarz, Uysal, Layer, Honegger, Wuttke, Koch and Zeck.
Figures
References
-
- Agnati L. F., Fuxe K., Zoli M., Ozini I., Toffano G., Ferraguti F. (1986). A correlation analysis of the regional distribution of central enkephalin and β-endorphin immunoreactive terminals and of opiate receptors in adult and old male rats. Evidence for the existence of two main types of communication in the central nervous sys. Acta Physiol. Scand. 128 201–207. 10.1111/j.1748-1716.1986.tb07967.x - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources