

Intrinsic and Synaptic Properties Shaping Diverse Behaviors of Neural Dynamics
- PMID: 32372936
- PMCID: PMC7187274
- DOI: 10.3389/fncom.2020.00026
Intrinsic and Synaptic Properties Shaping Diverse Behaviors of Neural Dynamics
Abstract
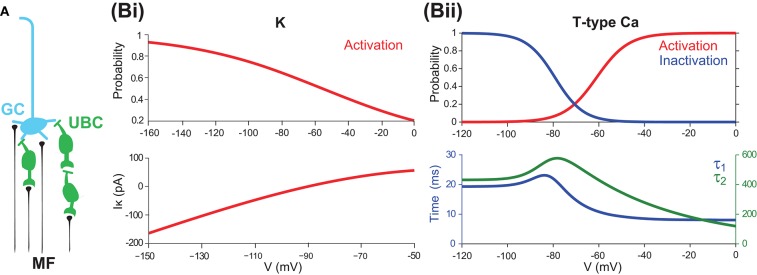
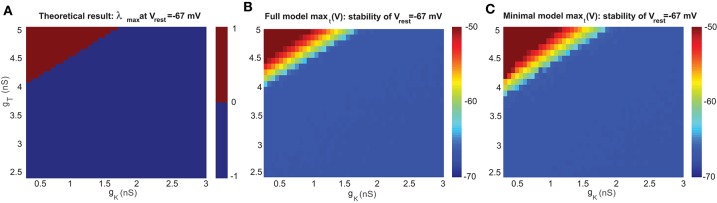
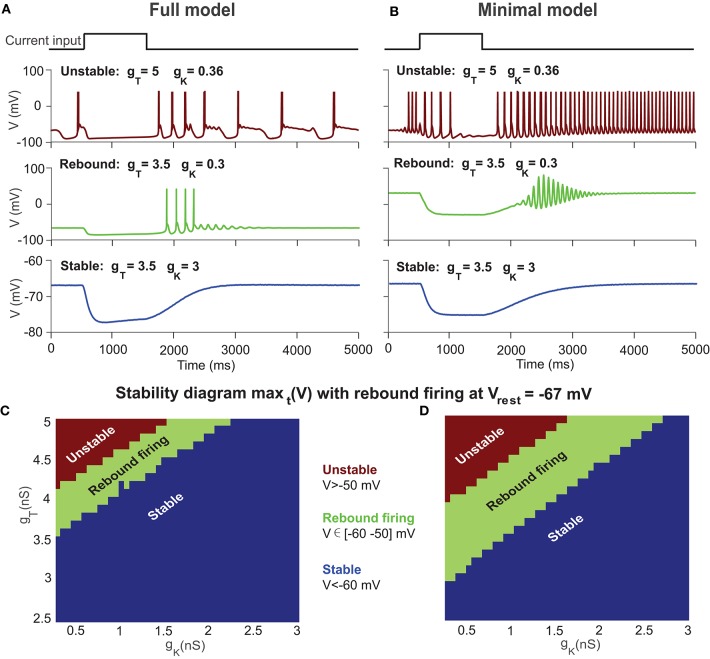
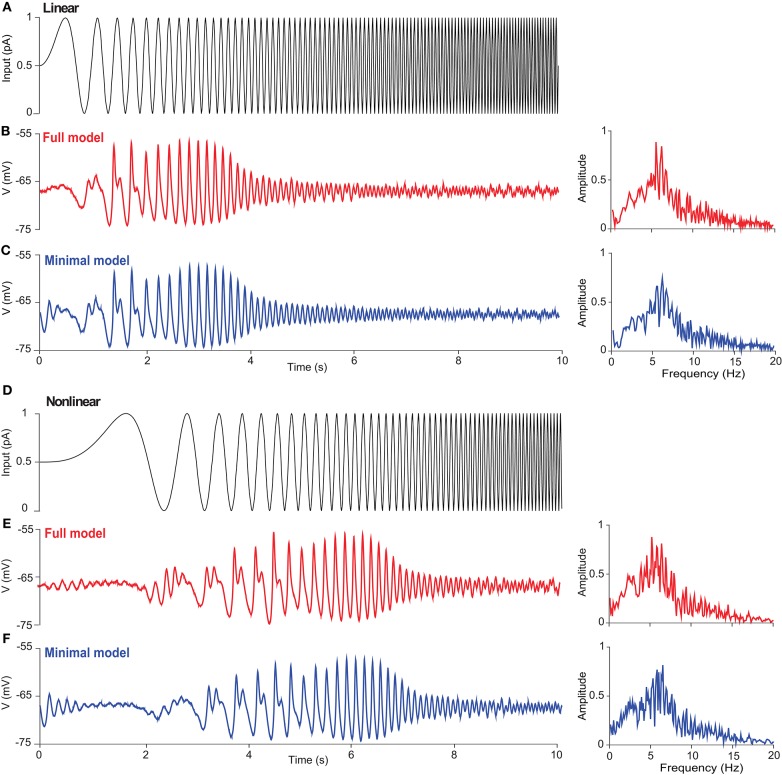
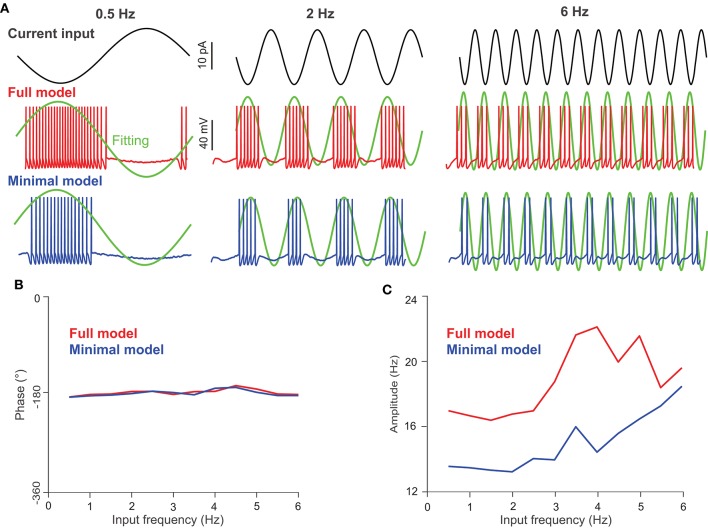
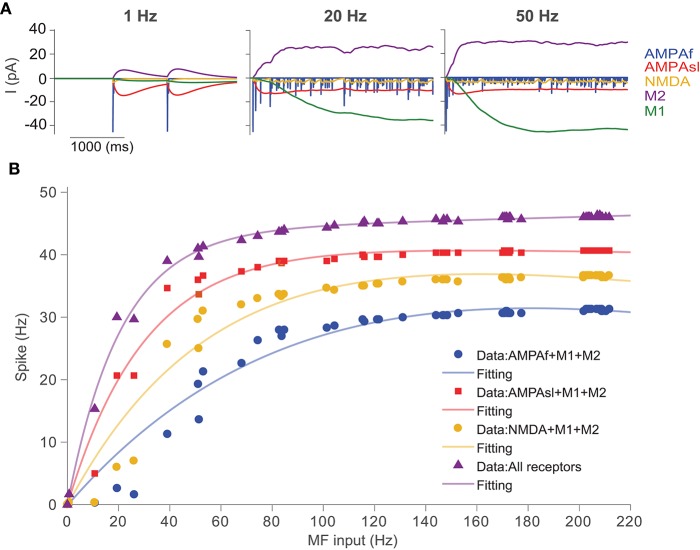
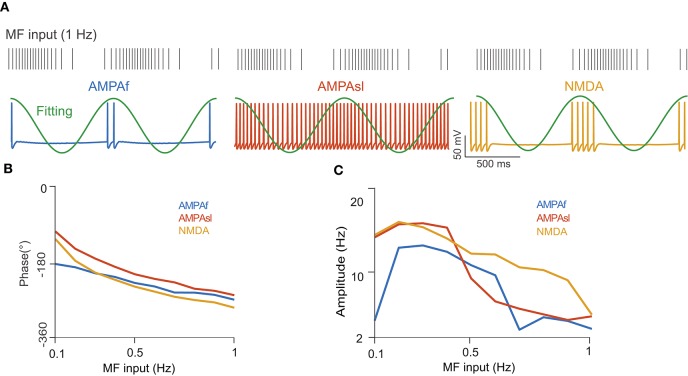
The majority of neurons in the neuronal system of the brain have a complex morphological structure, which diversifies the dynamics of neurons. In the granular layer of the cerebellum, there exists a unique cell type, the unipolar brush cell (UBC), that serves as an important relay cell for transferring information from outside mossy fibers to downstream granule cells. The distinguishing feature of the UBC is that it has a simple morphology, with only one short dendritic brush connected to its soma. Based on experimental evidence showing that UBCs exhibit a variety of dynamic behaviors, here we develop two simple models, one with a few detailed ion channels for simulation and the other one as a two-variable dynamical system for theoretical analysis, to characterize the intrinsic dynamics of UBCs. The reasonable values of the key channel parameters of the models can be determined by analysis of the stability of the resting membrane potential and the rebound firing properties of UBCs. Considered together with a large variety of synaptic dynamics installed on UBCs, we show that the simple-structured UBCs, as relay cells, can extend the range of dynamics and information from input mossy fibers to granule cells with low-frequency resonance and transfer stereotyped inputs to diverse amplitudes and phases of the output for downstream granule cells. These results suggest that neuronal computation, embedded within intrinsic ion channels and the diverse synaptic properties of single neurons without sophisticated morphology, can shape a large variety of dynamic behaviors to enhance the computational ability of local neuronal circuits.
Keywords: dynamical system; neural dynamic modeling; rebound firing; stability analysis; synaptic dynamics.
Copyright © 2020 An, Tang, Wang, Jia, Pei, Wang, Yu and Liu.
Figures
References
LinkOut - more resources
Full Text Sources