

Regulation of Energy Metabolism by Receptor Tyrosine Kinase Ligands
- PMID: 32372975
- PMCID: PMC7186430
- DOI: 10.3389/fphys.2020.00354
Regulation of Energy Metabolism by Receptor Tyrosine Kinase Ligands
Abstract
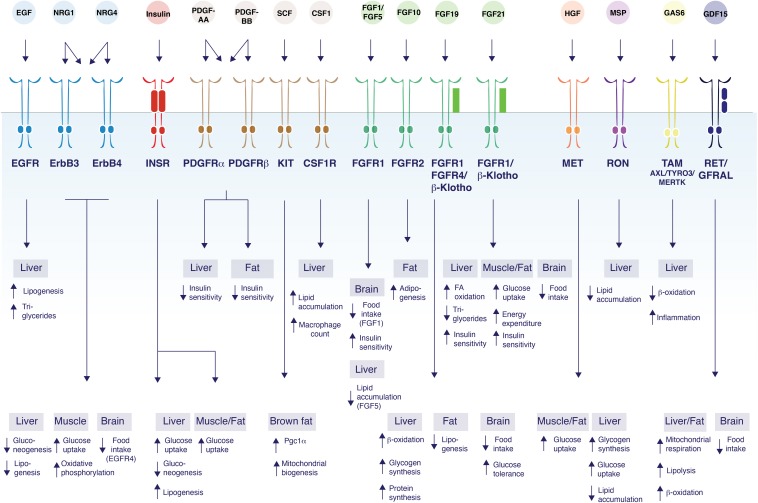
Metabolic diseases, such as diabetes, obesity, and fatty liver disease, have now reached epidemic proportions. Receptor tyrosine kinases (RTKs) are a family of cell surface receptors responding to growth factors, hormones, and cytokines to mediate a diverse set of fundamental cellular and metabolic signaling pathways. These ligands signal by endocrine, paracrine, or autocrine means in peripheral organs and in the central nervous system to control cellular and tissue-specific metabolic processes. Interestingly, the expression of many RTKs and their ligands are controlled by changes in metabolic demand, for example, during starvation, feeding, or obesity. In addition, studies of RTKs and their ligands in regulating energy homeostasis have revealed unexpected diversity in the mechanisms of action and their specific metabolic functions. Our current understanding of the molecular, biochemical and genetic control of energy homeostasis by the endocrine RTK ligands insulin, FGF21 and FGF19 are now relatively well understood. In addition to these classical endocrine signals, non-endocrine ligands can govern local energy regulation, and the intriguing crosstalk between the RTK family and the TGFβ receptor family demonstrates a signaling network that diversifies metabolic process between tissues. Thus, there is a need to increase our molecular and mechanistic understanding of signal diversification of RTK actions in metabolic disease. Here we review the known and emerging molecular mechanisms of RTK signaling that regulate systemic glucose and lipid metabolism, as well as highlighting unexpected roles of non-classical RTK ligands that crosstalk with other receptor pathways.
Keywords: glucose; lipids; metabolism; receptor tyrosine kinases; signaling.
Copyright © 2020 Zhao, Jung, Jiang and Svensson.
Figures
Similar articles
-
Minireview: Roles of Fibroblast Growth Factors 19 and 21 in Metabolic Regulation and Chronic Diseases.Mol Endocrinol. 2015 Oct;29(10):1400-13. doi: 10.1210/me.2015-1155. Epub 2015 Aug 26. Mol Endocrinol. 2015. PMID: 26308386 Free PMC article. Review.
-
Evaluating the landscape of gene cooperativity with receptor tyrosine kinases in liver tumorigenesis using transposon-mediated mutagenesis.J Hepatol. 2019 Mar;70(3):470-482. doi: 10.1016/j.jhep.2018.11.027. Epub 2018 Dec 6. J Hepatol. 2019. PMID: 30529386
-
Crosstalk coregulation mechanisms of G protein-coupled receptors and receptor tyrosine kinases.Methods Mol Biol. 2006;332:51-77. doi: 10.1385/1-59745-048-0:51. Methods Mol Biol. 2006. PMID: 16878685 Review.
-
The FGF metabolic axis.Front Med. 2019 Oct;13(5):511-530. doi: 10.1007/s11684-019-0711-y. Epub 2019 Sep 7. Front Med. 2019. PMID: 31495905 Free PMC article. Review.
-
Galectins as modulators of receptor tyrosine kinases signaling in health and disease.Cytokine Growth Factor Rev. 2021 Aug;60:89-106. doi: 10.1016/j.cytogfr.2021.03.004. Epub 2021 Mar 27. Cytokine Growth Factor Rev. 2021. PMID: 33863623 Review.
Cited by
-
AlphaFold2 enables accurate deorphanization of ligands to single-pass receptors.bioRxiv [Preprint]. 2023 Dec 15:2023.03.16.531341. doi: 10.1101/2023.03.16.531341. bioRxiv. 2023. Update in: Cell Syst. 2024 Nov 20;15(11):1046-1060.e3. doi: 10.1016/j.cels.2024.10.004. PMID: 36993313 Free PMC article. Updated. Preprint.
-
Pyrimidine: A Privileged Scaffold for the Development of Anticancer Agents as Protein Kinase Inhibitors (Recent Update).Curr Pharm Des. 2025;31(14):1100-1129. doi: 10.2174/0113816128346900241111115125. Curr Pharm Des. 2025. PMID: 39773052 Review.
-
Phosphoproteomic mapping reveals distinct signaling actions and activation of muscle protein synthesis by Isthmin-1.Elife. 2022 Sep 28;11:e80014. doi: 10.7554/eLife.80014. Elife. 2022. PMID: 36169399 Free PMC article.
-
Impact of obesity on the molecular response to a single bout of exercise in a preliminary human cohort.Obesity (Silver Spring). 2022 May;30(5):1091-1104. doi: 10.1002/oby.23419. Obesity (Silver Spring). 2022. PMID: 35470975 Free PMC article.
-
MicroRNA profiling of subcutaneous adipose tissue in periparturient dairy cows at high or moderate body condition.Sci Rep. 2022 Aug 30;12(1):14748. doi: 10.1038/s41598-022-18956-5. Sci Rep. 2022. PMID: 36042230 Free PMC article.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources