

DivIVA Controls Progeny Morphology and Diverse ParA Proteins Regulate Cell Division or Gliding Motility in Bdellovibrio bacteriovorus
- PMID: 32373080
- PMCID: PMC7186360
- DOI: 10.3389/fmicb.2020.00542
DivIVA Controls Progeny Morphology and Diverse ParA Proteins Regulate Cell Division or Gliding Motility in Bdellovibrio bacteriovorus
Abstract
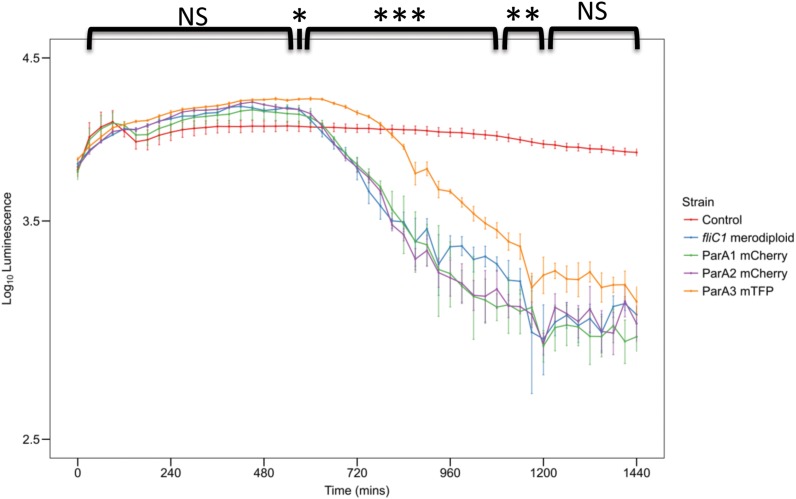
The predatory bacterium B. bacteriovorus grows and divides inside the periplasm of Gram-negative bacteria, forming a structure known as a bdelloplast. Cell division of predators inside the dead prey cell is not by binary fission but instead by synchronous division of a single elongated filamentous cell into odd or even numbers of progeny cells. Bdellovibrio replication and cell division processes are dependent on the finite level of nutrients available from inside the prey bacterium. The filamentous growth and division process of the predator maximizes the number of progeny produced by the finite nutrients in a way that binary fission could not. To learn more about such an unusual growth profile, we studied the role of DivIVA in the growing Bdellovibrio cell. This protein is well known for its link to polar cell growth and spore formation in Gram-positive bacteria, but little is known about its function in a predatory growth context. We show that DivIVA is expressed in the growing B. bacteriovorus cell and controls cell morphology during filamentous cell division, but not the number of progeny produced. Bacterial Two Hybrid (BTH) analysis shows DivIVA may interact with proteins that respond to metabolic indicators of amino-acid biosynthesis or changes in redox state. Such changes may be relevant signals to the predator, indicating the consumption of prey nutrients within the sealed bdelloplast environment. ParA, a chromosome segregation protein, also contributes to bacterial septation in many species. The B. bacteriovorus genome contains three ParA homologs; we identify a canonical ParAB pair required for predatory cell division and show a BTH interaction between a gene product encoded from the same operon as DivIVA with the canonical ParA. The remaining ParA proteins are both expressed in Bdellovibrio but are not required for predator cell division. Instead, one of these ParA proteins coordinates gliding motility, changing the frequency at which the cells reverse direction. Our work will prime further studies into how one bacterium can co-ordinate its cell division with the destruction of another bacterium that it dwells within.
Keywords: Bdellovibrio; DivIVA; ParAB; cell-morphology; predatory bacteria; septation.
Copyright © 2020 Milner, Ray, Saxon, Lambert, Till, Fenton and Sockett.
Figures







Similar articles
-
Binary or Nonbinary Fission? Reproductive Mode of a Predatory Bacterium Depends on Prey Size.mBio. 2023 Jun 27;14(3):e0077223. doi: 10.1128/mbio.00772-23. Epub 2023 May 10. mBio. 2023. PMID: 37162334 Free PMC article.
-
Distinct dynamics and proximity networks of hub proteins at the prey-invading cell pole in a predatory bacterium.J Bacteriol. 2024 Apr 18;206(4):e0001424. doi: 10.1128/jb.00014-24. Epub 2024 Mar 12. J Bacteriol. 2024. PMID: 38470120 Free PMC article.
-
Shadowing the actions of a predator: backlit fluorescent microscopy reveals synchronous nonbinary septation of predatory Bdellovibrio inside prey and exit through discrete bdelloplast pores.J Bacteriol. 2010 Dec;192(24):6329-35. doi: 10.1128/JB.00914-10. Epub 2010 Oct 8. J Bacteriol. 2010. PMID: 20935099 Free PMC article.
-
Chromosome structure and DNA replication dynamics during the life cycle of the predatory bacterium Bdellovibrio bacteriovorus.FEMS Microbiol Rev. 2023 Nov 1;47(6):fuad057. doi: 10.1093/femsre/fuad057. FEMS Microbiol Rev. 2023. PMID: 37791401 Free PMC article. Review.
-
Predatory lifestyle of Bdellovibrio bacteriovorus.Annu Rev Microbiol. 2009;63:523-39. doi: 10.1146/annurev.micro.091208.073346. Annu Rev Microbiol. 2009. PMID: 19575566 Review.
Cited by
-
Superficial incisional surgical site infections experimentally induced by Staphylococcus aureus in mice: the effect of Bdellovibrio bacteriovorus containing dressing.World J Microbiol Biotechnol. 2025 Jun 25;41(7):231. doi: 10.1007/s11274-025-04453-0. World J Microbiol Biotechnol. 2025. PMID: 40560484 Free PMC article.
-
Advances in cellular and molecular predatory biology of Bdellovibrio bacteriovorus six decades after discovery.Front Microbiol. 2023 May 15;14:1168709. doi: 10.3389/fmicb.2023.1168709. eCollection 2023. Front Microbiol. 2023. PMID: 37256055 Free PMC article. Review.
-
Binary or Nonbinary Fission? Reproductive Mode of a Predatory Bacterium Depends on Prey Size.mBio. 2023 Jun 27;14(3):e0077223. doi: 10.1128/mbio.00772-23. Epub 2023 May 10. mBio. 2023. PMID: 37162334 Free PMC article.
-
An optimized workflow to measure bacterial predation in microplates.STAR Protoc. 2022 Jan 20;3(1):101104. doi: 10.1016/j.xpro.2021.101104. eCollection 2022 Mar 18. STAR Protoc. 2022. PMID: 35098160 Free PMC article.
-
Chromosome choreography during the non-binary cell cycle of a predatory bacterium.Curr Biol. 2021 Sep 13;31(17):3707-3720.e5. doi: 10.1016/j.cub.2021.06.024. Epub 2021 Jul 12. Curr Biol. 2021. PMID: 34256020 Free PMC article.
References
-
- Altschul S. F., Gish W., Miller W., Myers E. W., Lipman D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215 403–410. - PubMed
-
- Ayala J. A., Garrido T., De Pedro M. A., Vicente M. (1994). Molecular Biology of Bacterial Septation. Bacterial Cell Wall. Amsterdam: Elsevier Science.
Grants and funding
LinkOut - more resources
Full Text Sources