

Entry of glucose- and glutamine-derived carbons into the citric acid cycle supports early steps of HIV-1 infection in CD4 T cells
- PMID: 32373781
- PMCID: PMC7199465
- DOI: 10.1038/s42255-019-0084-1
Entry of glucose- and glutamine-derived carbons into the citric acid cycle supports early steps of HIV-1 infection in CD4 T cells
Abstract
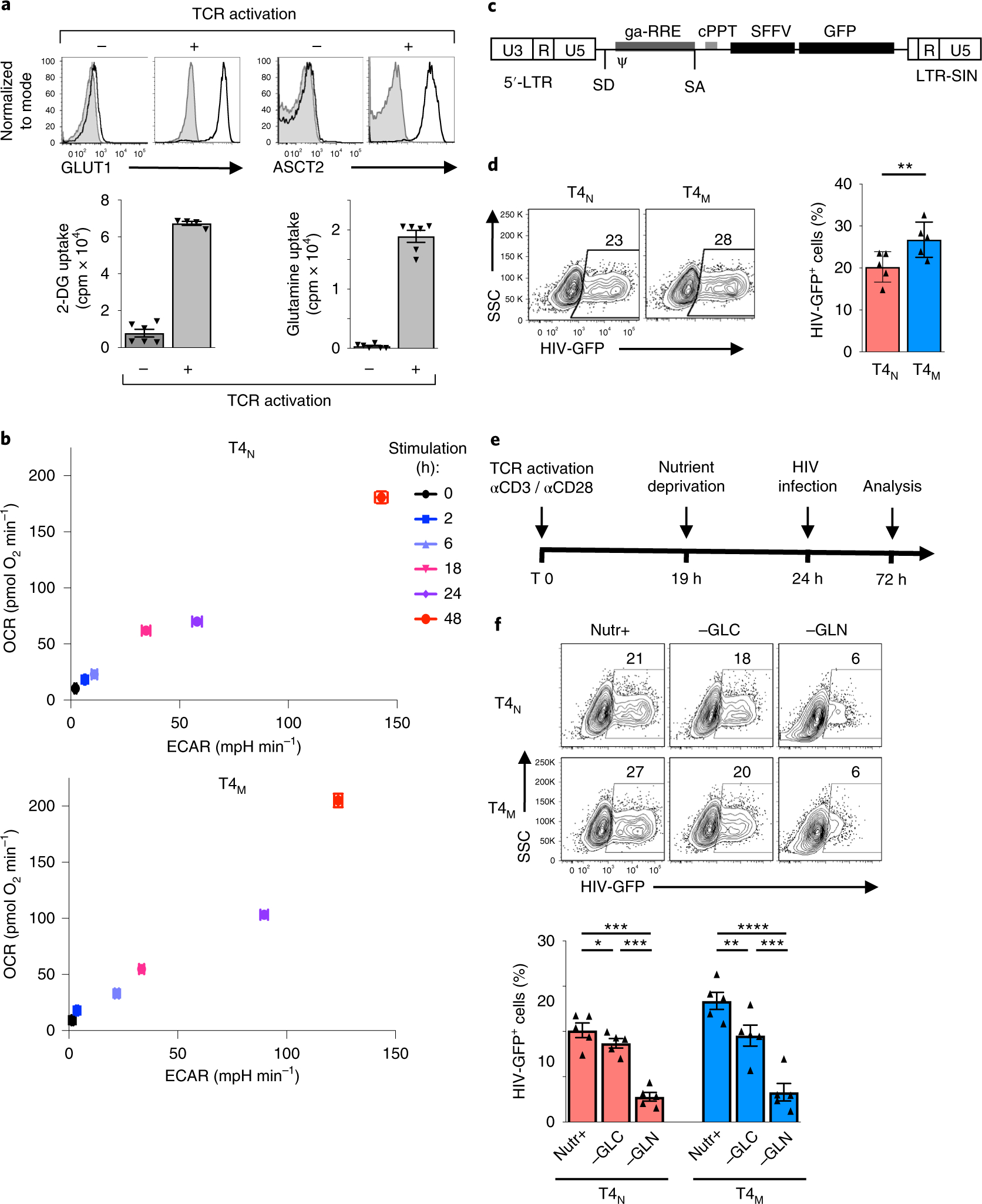
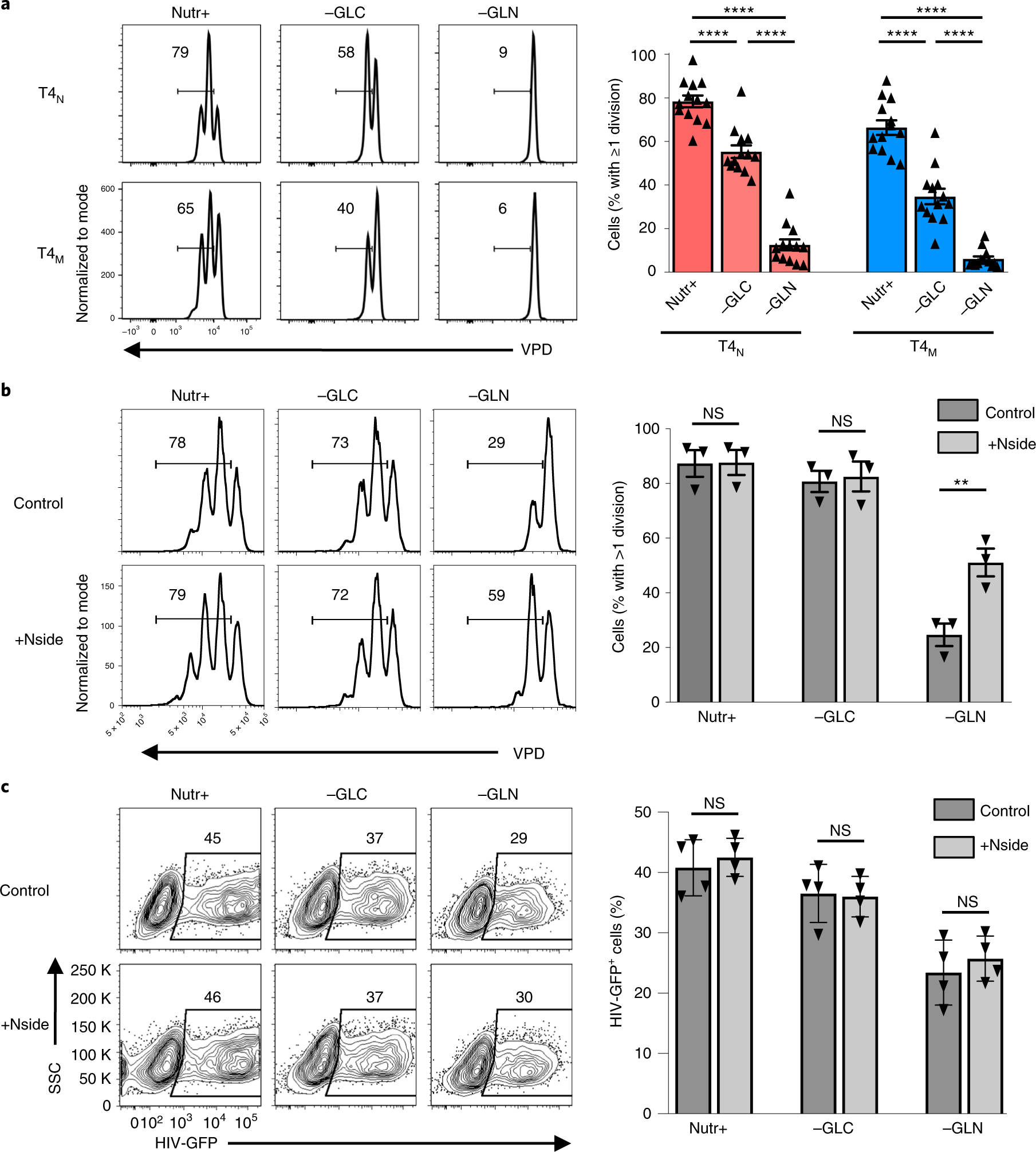
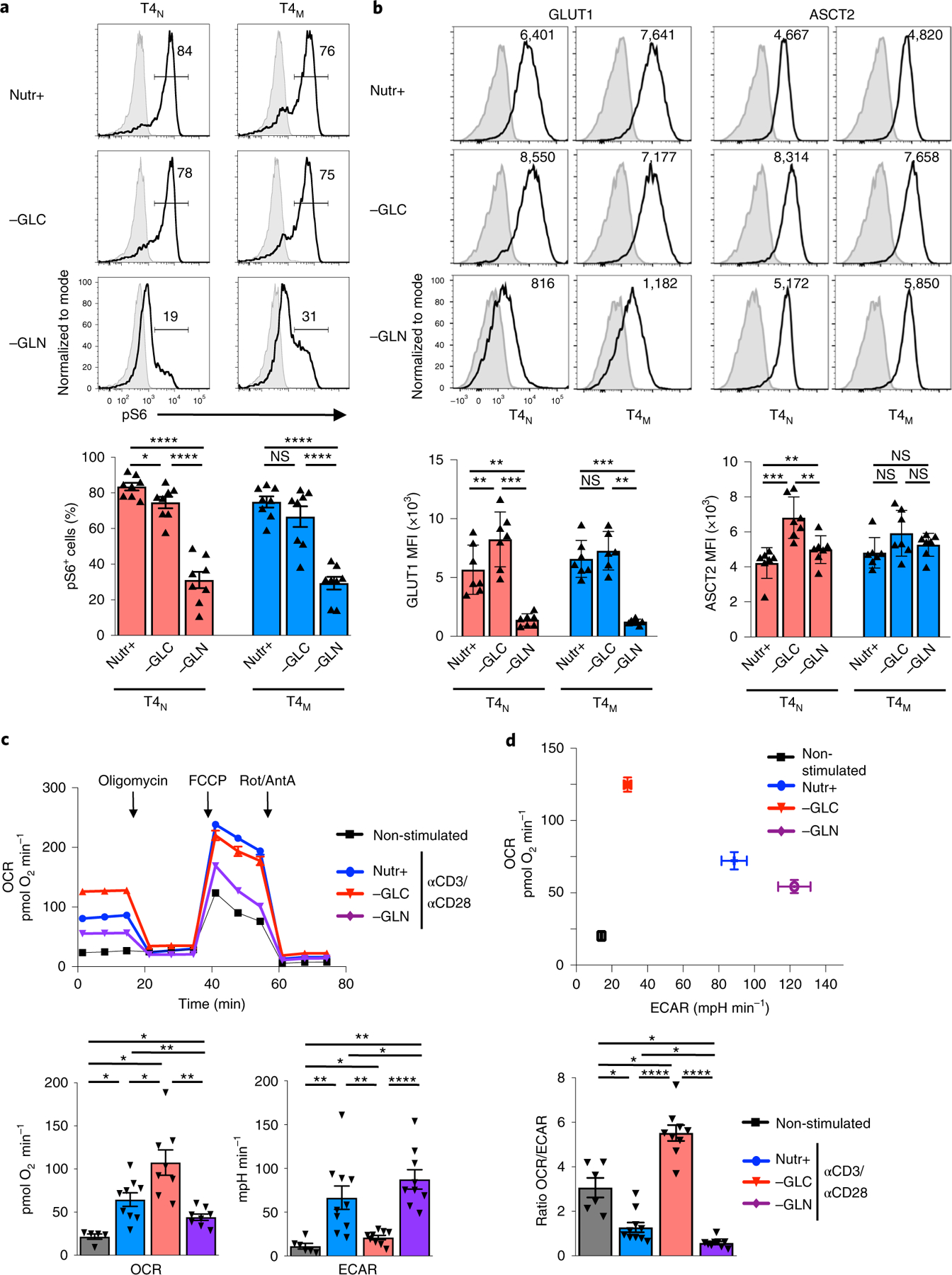
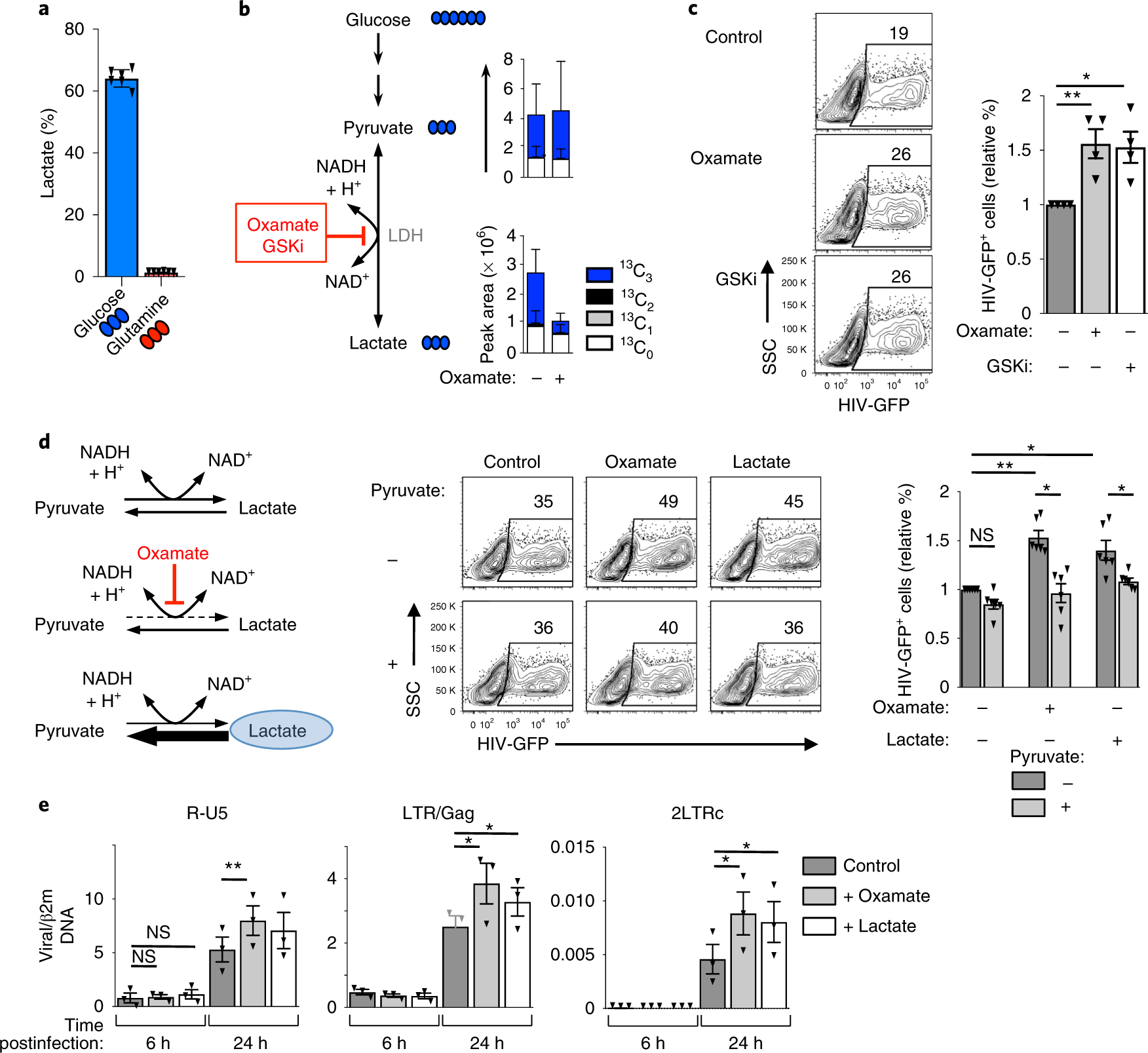
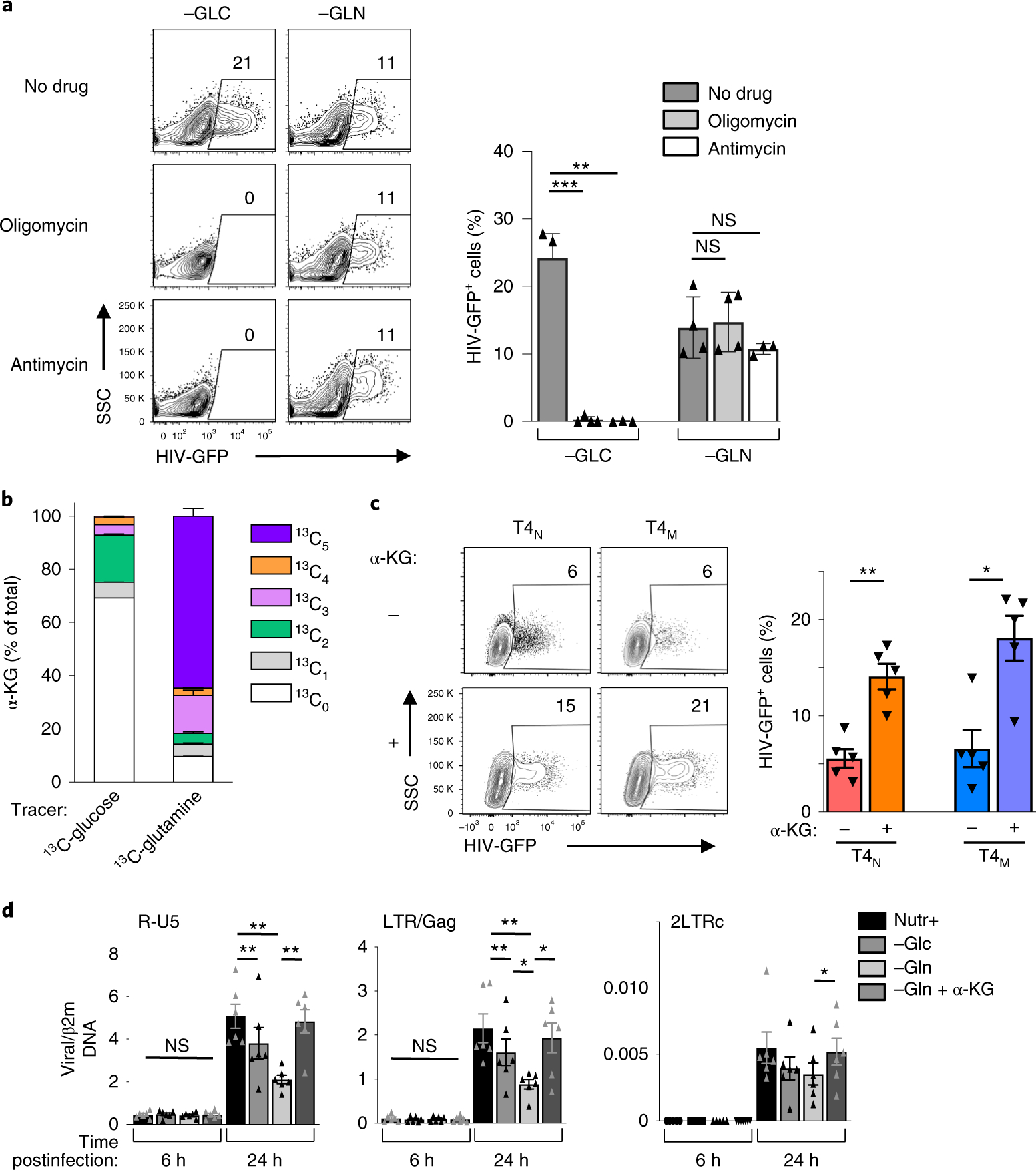
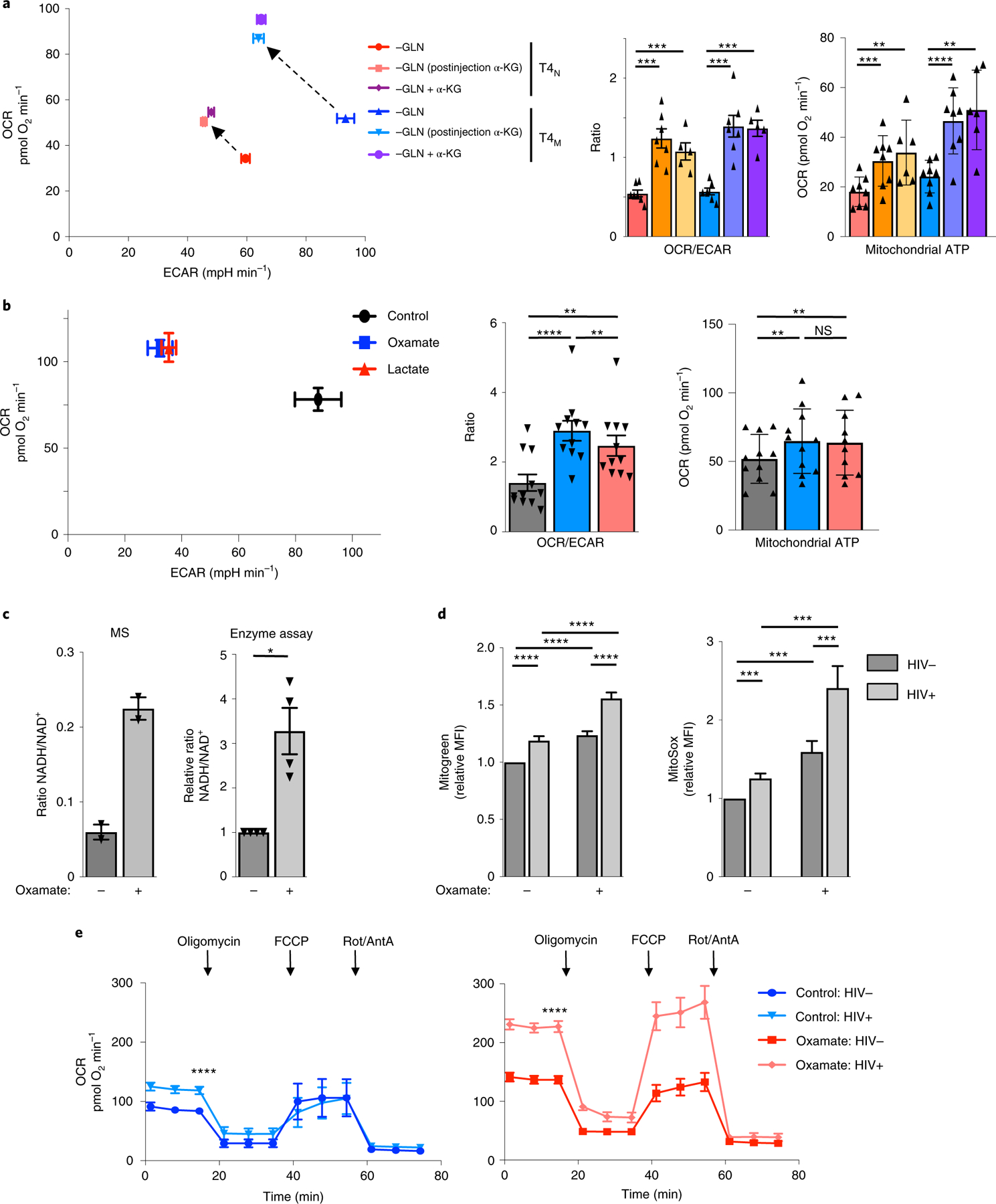
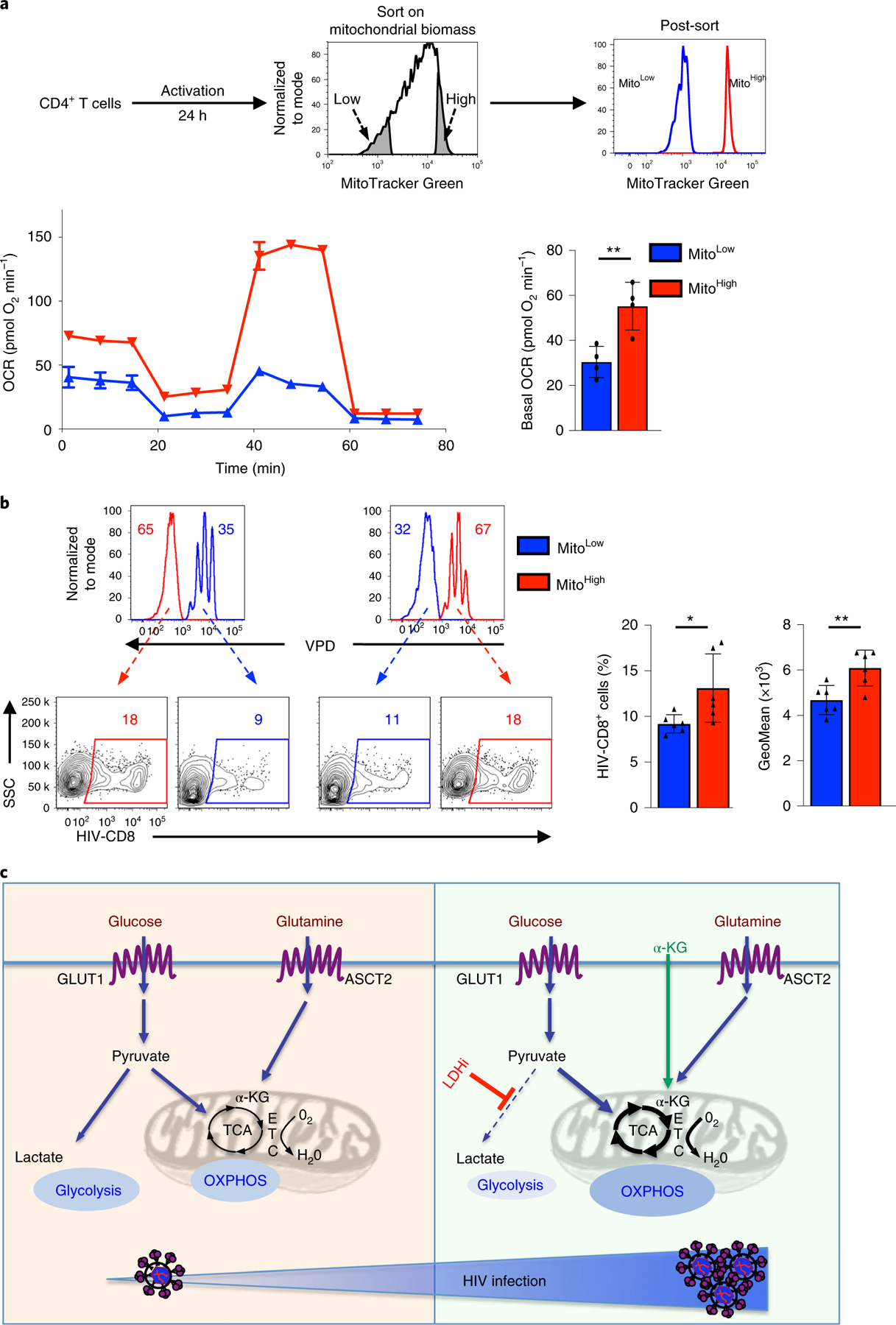
The susceptibility of CD4 T cells to human immunodeficiency virus 1 (HIV-1) infection is regulated by glucose and glutamine metabolism, but the relative contributions of these nutrients to infection are not known. Here we show that glutaminolysis is the major pathway fuelling the tricarboxylic acid (TCA) cycle and oxidative phosphorylation (OXPHOS) in T-cell receptor-stimulated naïve, as well as memory CD4, subsets and is required for optimal HIV-1 infection. Under conditions of attenuated glutaminolysis, the α-ketoglutarate (α-KG) TCA rescues early steps in infection; exogenous α-KG promotes HIV-1 reverse transcription, rendering both naïve and memory cells more sensitive to infection. Blocking the glycolytic flux of pyruvate to lactate results in altered glucose carbon allocation to TCA and pentose phosphate pathway intermediates, an increase in OXPHOS and augmented HIV-1 reverse transcription. Moreover, HIV-1 infection is significantly higher in CD4 T cells selected on the basis of high mitochondrial biomass and OXPHOS activity. Therefore, the OXPHOS/aerobic glycolysis balance is a major regulator of HIV-1 infection in CD4 T lymphocytes.
Conflict of interest statement
Competing interests M.S. and N.T. are inventors on patent WO2010079208. M.S., C.M. and N.T. are inventors on patent WO/2004/096841 and M.S. is an inventor on patent WO/2012/035369. All patents are owned by the CNRS and cover the use of RBD ligands for metabolite transporter detection. N.T. no longer owns any patent rights. M.S. is a co-founder of METAFORA Biosystems.
Figures
References
-
- Rathmell JC, Elstrom RL, Cinalli RM & Thompson CB Activated Akt promotes increased resting T cell size, CD28-independent T cell growth, and development of autoimmunity and lymphoma. Eur. J. Immunol 33, 2223–2232 (2003). - PubMed
-
- Frauwirth KA et al. The CD28 signaling pathway regulates glucose metabolism. Immunity 16, 769–777 (2002). - PubMed
-
- Manel N et al. The HTLV receptor is an early T-cell activation marker whose expression requires de novo protein synthesis. Blood 101, 1913–1918 (2003). - PubMed
-
- Curi R et al. Glutamine, gene expression, and cell function. Front. Biosci 12, 344–357 (2007). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
