

Proximity biotinylation reveals novel secreted dense granule proteins of Toxoplasma gondii bradyzoites
- PMID: 32374791
- PMCID: PMC7202600
- DOI: 10.1371/journal.pone.0232552
Proximity biotinylation reveals novel secreted dense granule proteins of Toxoplasma gondii bradyzoites
Abstract
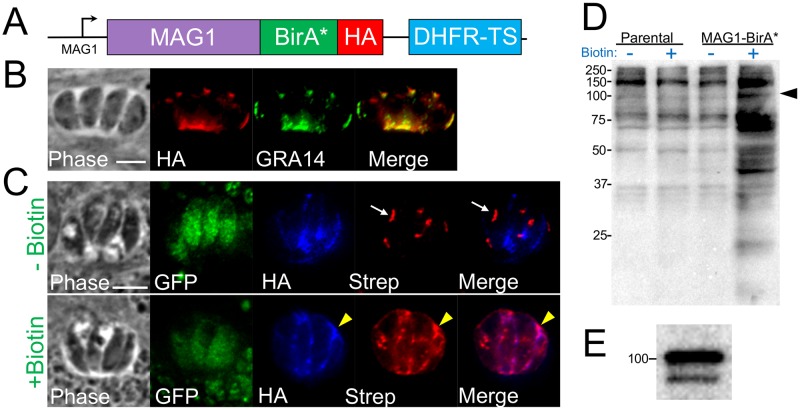
Toxoplasma gondii is an obligate intracellular parasite which is capable of establishing life-long chronic infection in any mammalian host. During the intracellular life cycle, the parasite secretes an array of proteins into the parasitophorous vacuole (PV) where it resides. Specialized organelles called the dense granules secrete GRA proteins that are known to participate in nutrient acquisition, immune evasion, and host cell-cycle manipulation. Although many GRAs have been discovered which are expressed during the acute infection mediated by tachyzoites, little is known about those that participate in the chronic infection mediated by the bradyzoite form of the parasite. In this study, we sought to uncover novel bradyzoite-upregulated GRA proteins using proximity biotinylation, which we previously used to examine the secreted proteome of the tachyzoites. Using a fusion of the bradyzoite upregulated protein MAG1 to BirA* as bait and a strain with improved switch efficiency, we identified a number of novel GRA proteins which are expressed in bradyzoites. After using the CRISPR/Cas9 system to characterize these proteins by gene knockout, we focused on one of these GRAs (GRA55) and found it was important for the establishment or maintenance of cysts in the mouse brain. These findings highlight new components of the GRA proteome of the tissue-cyst life stage of T. gondii and identify potential targets that are important for maintenance of parasite persistence in vivo.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



Similar articles
-
Functional Characterization of 15 Novel Dense Granule Proteins in Toxoplasma gondii Using the CRISPR-Cas9 System.Microbiol Spectr. 2023 Feb 14;11(1):e0307822. doi: 10.1128/spectrum.03078-22. Epub 2022 Dec 14. Microbiol Spectr. 2023. PMID: 36515555 Free PMC article.
-
In Vivo Biotinylation of the Toxoplasma Parasitophorous Vacuole Reveals Novel Dense Granule Proteins Important for Parasite Growth and Pathogenesis.mBio. 2016 Aug 2;7(4):e00808-16. doi: 10.1128/mBio.00808-16. mBio. 2016. PMID: 27486190 Free PMC article.
-
Biotinylation of the Neospora caninum parasitophorous vacuole reveals novel dense granule proteins.Parasit Vectors. 2021 Oct 9;14(1):521. doi: 10.1186/s13071-021-05023-7. Parasit Vectors. 2021. PMID: 34625097 Free PMC article.
-
GRA proteins of Toxoplasma gondii: maintenance of host-parasite interactions across the parasitophorous vacuolar membrane.Korean J Parasitol. 2009 Oct;47 Suppl(Suppl):S29-37. doi: 10.3347/kjp.2009.47.S.S29. Korean J Parasitol. 2009. PMID: 19885333 Free PMC article. Review.
-
Communication between Toxoplasma gondii and its host: impact on parasite growth, development, immune evasion, and virulence.APMIS. 2009 May;117(5-6):458-76. doi: 10.1111/j.1600-0463.2009.02453.x. APMIS. 2009. PMID: 19400868 Free PMC article. Review.
Cited by
-
Development and validation of a machine learning algorithm prediction for dense granule proteins in Apicomplexa.Parasit Vectors. 2023 Mar 14;16(1):98. doi: 10.1186/s13071-023-05698-0. Parasit Vectors. 2023. PMID: 36918932 Free PMC article.
-
Systematic characterization of all Toxoplasma gondii TBC domain-containing proteins identifies an essential regulator of Rab2 in the secretory pathway.PLoS Biol. 2024 May 7;22(5):e3002634. doi: 10.1371/journal.pbio.3002634. eCollection 2024 May. PLoS Biol. 2024. PMID: 38713739 Free PMC article.
-
Toxoplasma gondii Matrix Antigen 1 Is a Secreted Immunomodulatory Effector.mBio. 2021 May 18;12(3):e00603-21. doi: 10.1128/mBio.00603-21. mBio. 2021. PMID: 34006649 Free PMC article.
-
Toxoplasma bradyzoites exhibit physiological plasticity of calcium and energy stores controlling motility and egress.Elife. 2021 Dec 3;10:e73011. doi: 10.7554/eLife.73011. Elife. 2021. PMID: 34860156 Free PMC article.
-
Efficient Gene Knockout and Knockdown Systems in Neospora caninum Enable Rapid Discovery and Functional Assessment of Novel Proteins.mSphere. 2022 Feb 23;7(1):e0089621. doi: 10.1128/msphere.00896-21. Epub 2022 Jan 12. mSphere. 2022. PMID: 35019667 Free PMC article.
