

eIF4G has intrinsic G-quadruplex binding activity that is required for tiRNA function
- PMID: 32374873
- PMCID: PMC7293036
- DOI: 10.1093/nar/gkaa336
eIF4G has intrinsic G-quadruplex binding activity that is required for tiRNA function
Abstract
As cells encounter adverse environmental conditions, such as hypoxia, oxidative stress or nutrient deprivation, they trigger stress response pathways to protect themselves until transient stresses have passed. Inhibition of translation is a key component of such cellular stress responses and mounting evidence has revealed the importance of a class of tRNA-derived small RNAs called tiRNAs in this process. The most potent of these small RNAs are those with the capability of assembling into tetrameric G-quadruplex (G4) structures. However, the mechanism by which these small RNAs inhibit translation has yet to be elucidated. Here we show that eIF4G, the major scaffolding protein in the translation initiation complex, directly binds G4s and this activity is required for tiRNA-mediated translation repression. Targeting of eIF4G results in an impairment of 40S ribosome scanning on mRNAs leading to the formation of eIF2α-independent stress granules. Our data reveals the mechanism by which tiRNAs inhibit translation and demonstrates novel activity for eIF4G in the regulation of translation.
© The Author(s) 2020. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
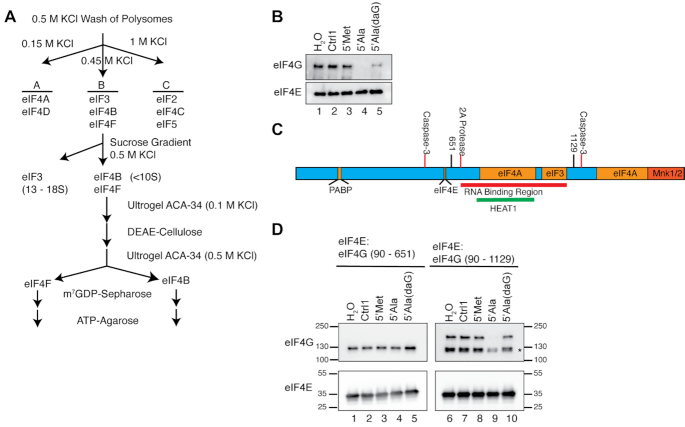
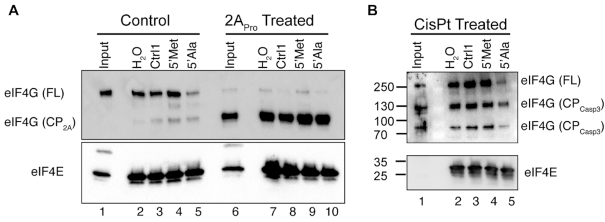
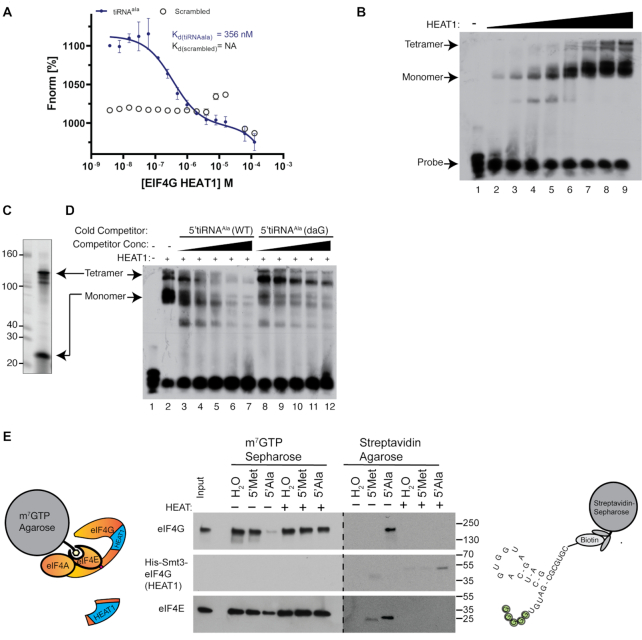
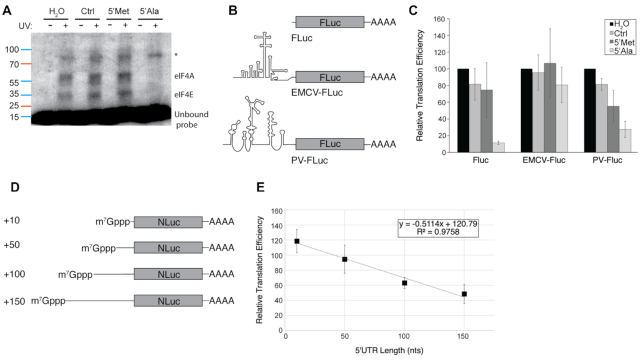
References
-
- Holcik M., Sonenberg N.. Translational control in stress and apoptosis. Nat. Rev. Mol. Cell Biol. 2005; 6:318–327. - PubMed
-
- Akiyama Y., Lyons S.M., Fay M.M., Abe T., Anderson P., Ivanov P.. Multiple ribonuclease A family members cleave transfer RNAs in response to stress. 2019; bioRxiv doi:21 October 2019, preprint: not peer reviewed 10.1101/811174. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
