

Androgen attenuates the inactivating phospho-Ser-127 modification of yes-associated protein 1 (YAP1) and promotes YAP1 nuclear abundance and activity
- PMID: 32376689
- PMCID: PMC7307196
- DOI: 10.1074/jbc.RA120.013794
Androgen attenuates the inactivating phospho-Ser-127 modification of yes-associated protein 1 (YAP1) and promotes YAP1 nuclear abundance and activity
Abstract
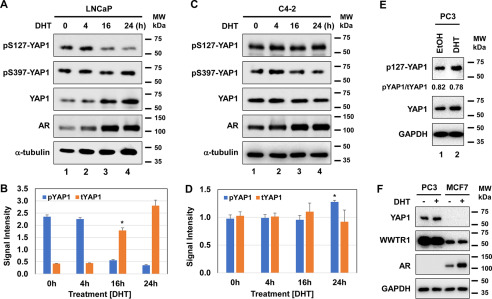
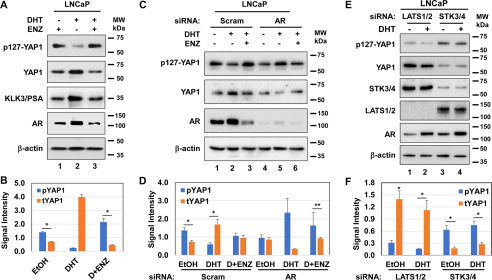
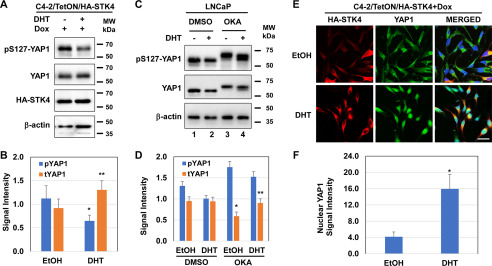
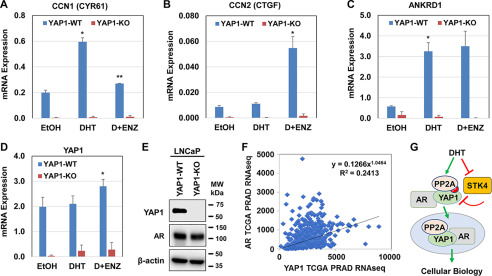
The transcriptional coactivator YAP1 (yes-associated protein 1) regulates cell proliferation, cell-cell interactions, organ size, and tumorigenesis. Post-transcriptional modifications and nuclear translocation of YAP1 are crucial for its nuclear activity. The objective of this study was to elucidate the mechanism by which the steroid hormone androgen regulates YAP1 nuclear entry and functions in several human prostate cancer cell lines. We demonstrate that androgen exposure suppresses the inactivating post-translational modification phospho-Ser-127 in YAP1, coinciding with increased YAP1 nuclear accumulation and activity. Pharmacological and genetic experiments revealed that intact androgen receptor signaling is necessary for androgen's inactivating effect on phospho-Ser-127 levels and increased YAP1 nuclear entry. We also found that androgen exposure antagonizes Ser/Thr kinase 4 (STK4/MST1) signaling, stimulates the activity of protein phosphatase 2A, and thereby attenuates the phospho-Ser-127 modification and promotes YAP1 nuclear localization. Results from quantitative RT-PCR and CRISPR/Cas9-aided gene knockout experiments indicated that androgen differentially regulates YAP1-dependent gene expression. Furthermore, an unbiased computational analysis of the prostate cancer data from The Cancer Genome Atlas revealed that YAP1 and androgen receptor transcript levels correlate with each other in prostate cancer tissues. These findings indicate that androgen regulates YAP1 nuclear localization and its transcriptional activity through the androgen receptor-STK4/MST1-protein phosphatase 2A axis, which may have important implications for human diseases such as prostate cancer.
Keywords: Hippo pathway; Hippo/MST1/STK4; YAP1; YAP1 nuclear localization; androgen; androgen receptor; androgen/AR signaling; androgens; cell signaling; gene transcription; nuclear translocation; phosphorylation; post-translational modification; protein phosphatase PP2A; protein phosphorylation; protein serine/threonine phosphatase (PSP); protein–protein interaction; signal transduction; yes-associated protein (YAP).
© 2020 Cinar et al.
Conflict of interest statement
Conflict of interest—The authors declare no competing conflicts of interest with the content of this article.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
