

Extracellular Vesicles from Aspergillus flavus Induce M1 Polarization In Vitro
- PMID: 32376699
- PMCID: PMC7203453
- DOI: 10.1128/mSphere.00190-20
Extracellular Vesicles from Aspergillus flavus Induce M1 Polarization In Vitro
Abstract
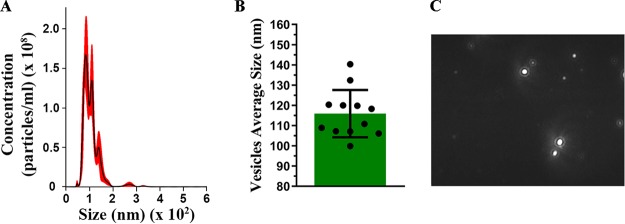
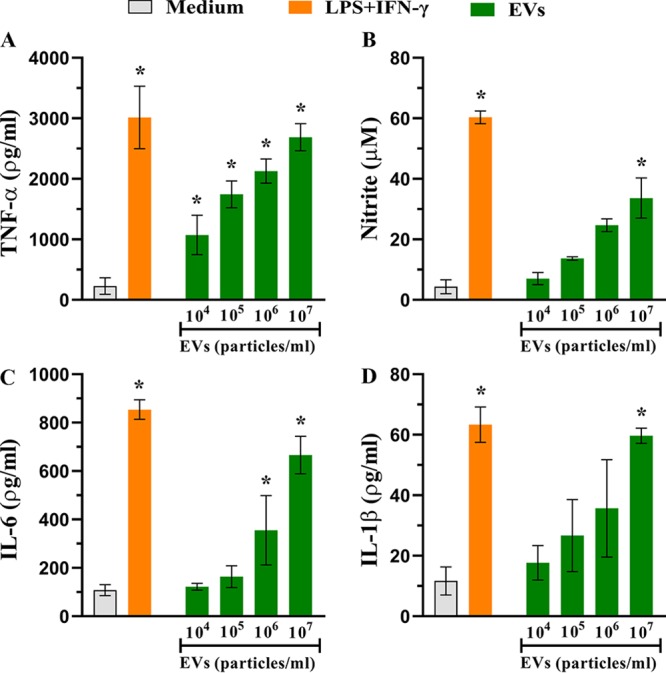
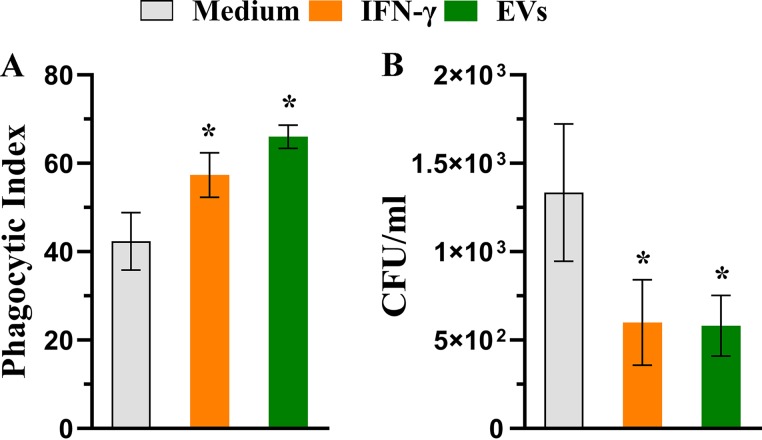
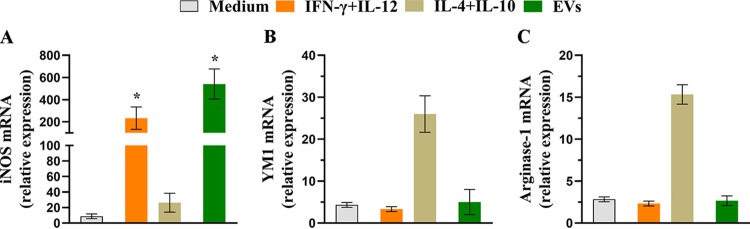
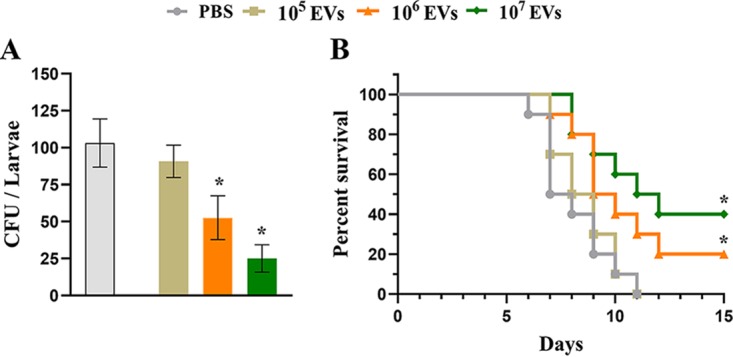
Aspergillus flavus, a ubiquitous and saprophytic fungus, is the second most common cause of aspergillosis worldwide. Several mechanisms contribute to the establishment of the fungal infection. Extracellular vesicles (EVs) have been described as "virulence factor delivery bags" in several fungal species, demonstrating a crucial role during the infection. In this study, we evaluated production of A. flavus EVs and their immunomodulatory functions. We verified that A. flavus EVs induce macrophages to produce inflammatory mediators, such as nitric oxide, tumor necrosis factor alpha (TNF-α), interleukin-6 (IL-6), and IL-1β. Furthermore, the A. flavus EVs enhance phagocytosis and killing by macrophages and induce M1 macrophage polarization in vitro In addition, a prior inoculation of A. flavus EVs in Galleria mellonella larvae resulted in a protective effect against the fungal infection. Our findings suggest that A. flavus EVs are biologically active and affect the interaction between A. flavus and host immune cells, priming the innate immune system to eliminate the fungal infection. Collectively, our results suggest that A. flavus EVs play a crucial role in aspergillosis.IMPORTANCE Immunocompromised patients are susceptible to several fungal infections. The genus Aspergillus can cause increased morbidity and mortality. Developing new therapies is essential to understand the fungal biology mechanisms. Fungal EVs carry important virulence factors, thus playing pivotal roles in fungal pathophysiology. No study to date has reported EV production by Aspergillus flavus, a fungus considered to be the second most common cause of aspergillosis and relevant food contaminator found worldwide. In this study, we produced A. flavus EVs and evaluated the in vitro immunomodulatory effects of EVs on bone marrow-derived macrophages (BMDMs) and in vivo effects in a Galleria mellonella model.
Keywords: Aspergillus flavus; Galleria mellonella; cytokine; extracellular vesicles; killing; macrophage; phagocytosis; polarization.
Copyright © 2020 Brauer et al.
Figures
References
-
- Jackson SA, Dobson A. 2016. Yeasts and molds: Aspergillus flavus. Reference module in food science. Elsevier, Amsterdam, Netherlands. doi: 10.1016/B978-0-08-100596-5.01086-6. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
