

Chemical activation of SAT1 corrects diet-induced metabolic syndrome
- PMID: 32376874
- PMCID: PMC7494776
- DOI: 10.1038/s41418-020-0550-z
Chemical activation of SAT1 corrects diet-induced metabolic syndrome
Abstract
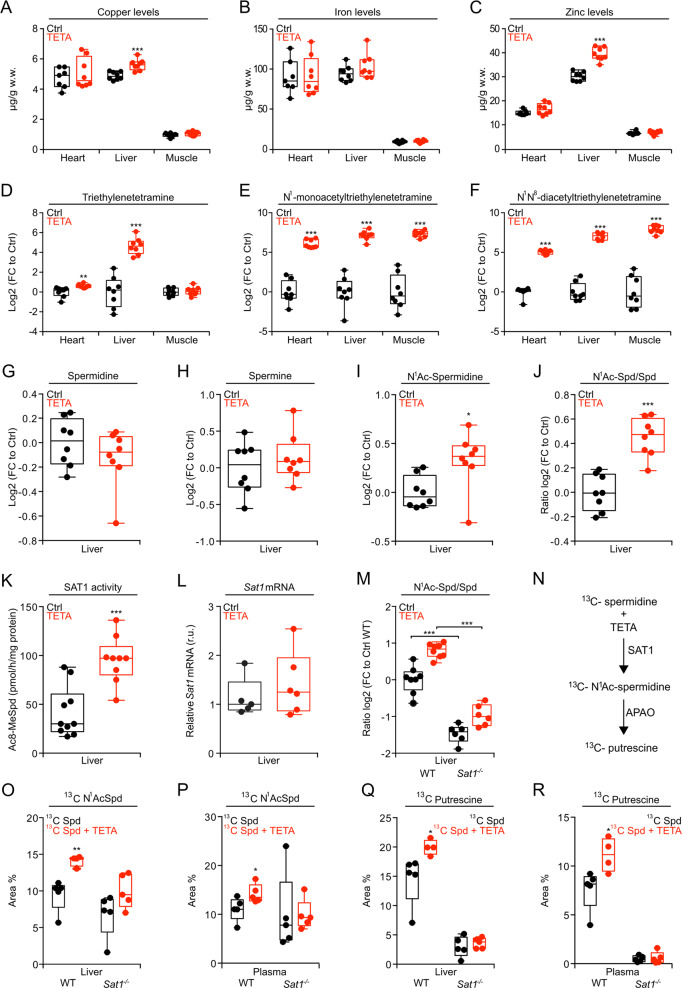
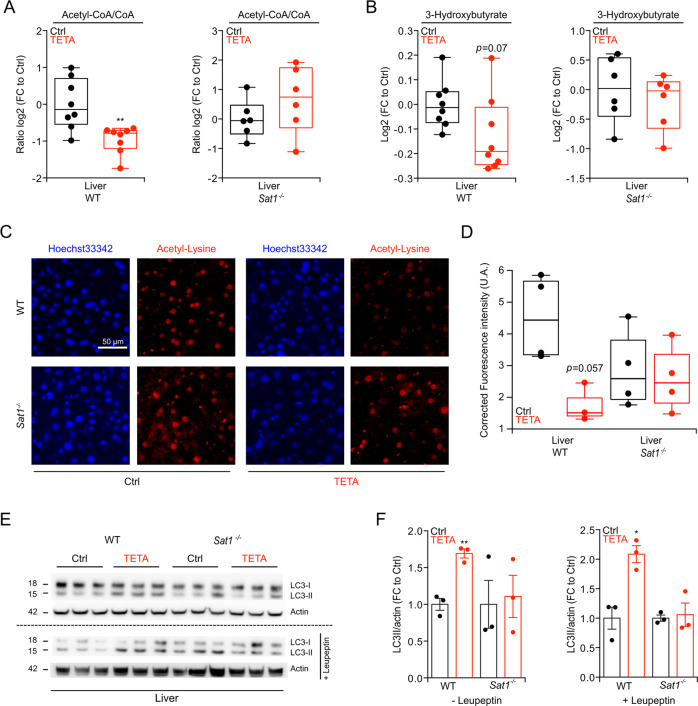
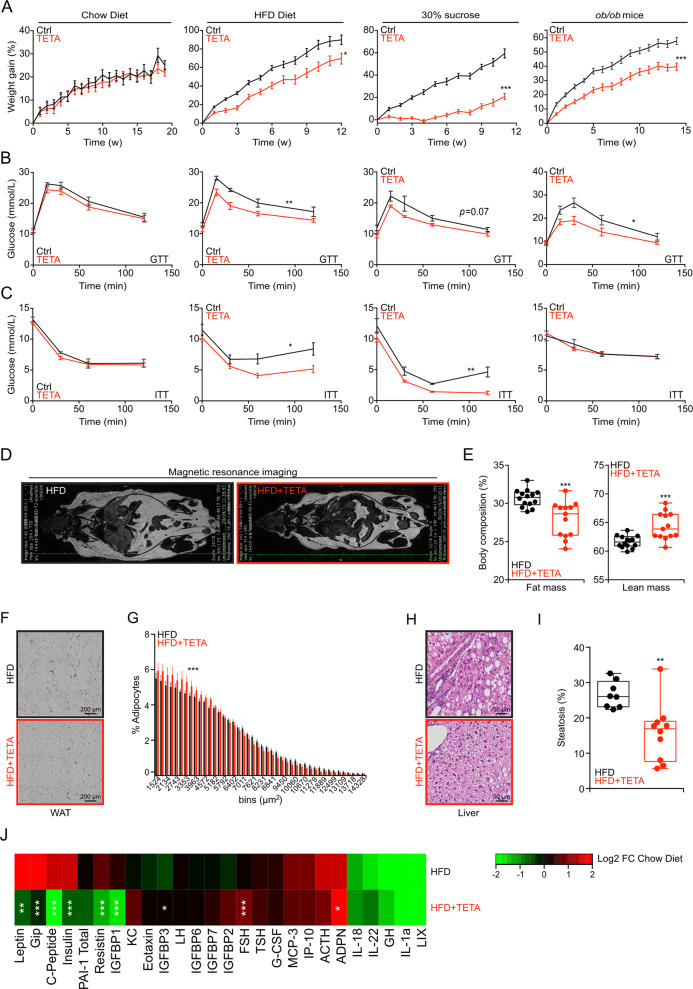
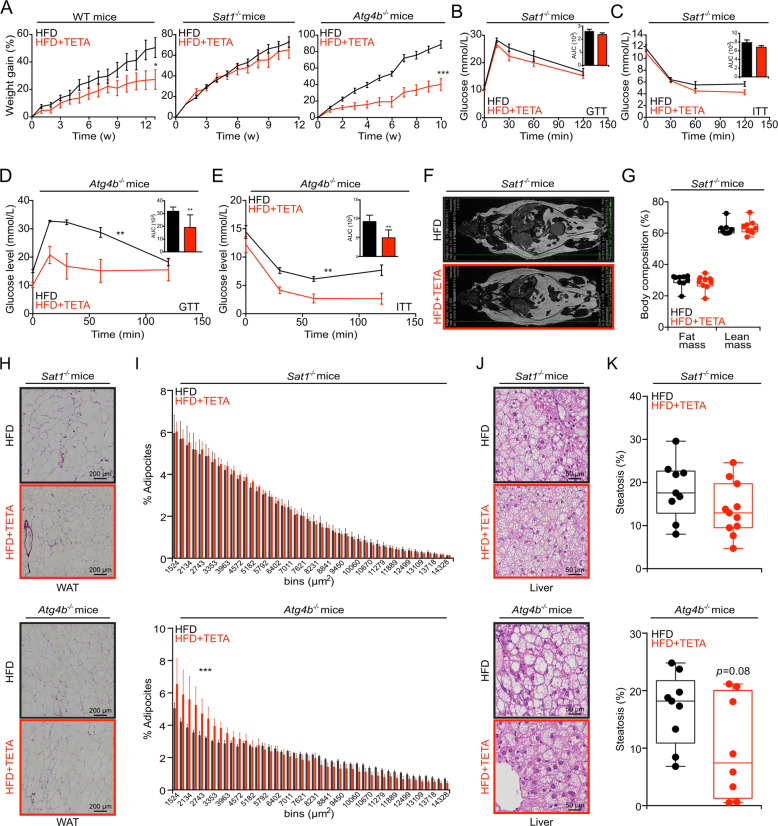
The pharmacological targeting of polyamine metabolism is currently under the spotlight for its potential in the prevention and treatment of several age-associated disorders. Here, we report the finding that triethylenetetramine dihydrochloride (TETA), a copper-chelator agent that can be safely administered to patients for the long-term treatment of Wilson disease, exerts therapeutic benefits in animals challenged with hypercaloric dietary regimens. TETA reduced obesity induced by high-fat diet, excessive sucrose intake, or leptin deficiency, as it reduced glucose intolerance and hepatosteatosis, but induced autophagy. Mechanistically, these effects did not involve the depletion of copper from plasma or internal organs. Rather, the TETA effects relied on the activation of an energy-consuming polyamine catabolism, secondary to the stabilization of spermidine/spermine N1-acetyltransferase-1 (SAT1) by TETA, resulting in enhanced enzymatic activity of SAT. All the positive effects of TETA on high-fat diet-induced metabolic syndrome were lost in SAT1-deficient mice. Altogether, these results suggest novel health-promoting effects of TETA that might be taken advantage of for the prevention or treatment of obesity.
Conflict of interest statement
GK and FM are co-founders of Samsara Therapeutics. GK and FP hold a patent protecting new medical use for TETA. FM is a co-founder of The Longevity Labs. TE has equity interests in The Longevity Labs. GK is a consultant for The Longevity Labs.
Figures
Similar articles
-
Triethylenetetramine (trientine): a caloric restriction mimetic with a new mode of action.Autophagy. 2020 Aug;16(8):1534-1536. doi: 10.1080/15548627.2020.1778293. Epub 2020 Jun 16. Autophagy. 2020. PMID: 32544364 Free PMC article.
-
Metabolism of triethylenetetramine and 1,12-diamino-3,6,9-triazadodecane by the spermidine/spermine-N(1)-acetyltransferase and thialysine acetyltransferase.Drug Metab Dispos. 2013 Jan;41(1):30-2. doi: 10.1124/dmd.112.047274. Epub 2012 Sep 28. Drug Metab Dispos. 2013. PMID: 23024204
-
Complex N-acetylation of triethylenetetramine.Drug Metab Dispos. 2011 Dec;39(12):2242-9. doi: 10.1124/dmd.111.041798. Epub 2011 Aug 30. Drug Metab Dispos. 2011. PMID: 21878558 Free PMC article.
-
Therapeutic potential of copper chelation with triethylenetetramine in managing diabetes mellitus and Alzheimer's disease.Drugs. 2011 Jul 9;71(10):1281-320. doi: 10.2165/11591370-000000000-00000. Drugs. 2011. PMID: 21770477 Review.
-
Triethylenetetramine pharmacology and its clinical applications.Mol Cancer Ther. 2010 Sep;9(9):2458-67. doi: 10.1158/1535-7163.MCT-10-0523. Epub 2010 Jul 26. Mol Cancer Ther. 2010. PMID: 20660601 Review.
Cited by
-
Current Status of Autophagy Enhancers in Metabolic Disorders and Other Diseases.Front Cell Dev Biol. 2022 Feb 14;10:811701. doi: 10.3389/fcell.2022.811701. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35237600 Free PMC article. Review.
-
Caloric restriction induced epigenetic effects on aging.Front Cell Dev Biol. 2023 Jan 13;10:1079920. doi: 10.3389/fcell.2022.1079920. eCollection 2022. Front Cell Dev Biol. 2023. PMID: 36712965 Free PMC article. Review.
-
The central role of creatine and polyamines in fetal growth restriction.FASEB J. 2024 Dec 15;38(23):e70222. doi: 10.1096/fj.202401946R. FASEB J. 2024. PMID: 39614665 Free PMC article.
-
Circulating acetylated polyamines correlate with Covid-19 severity in cancer patients.Aging (Albany NY). 2021 Sep 13;13(17):20860-20885. doi: 10.18632/aging.203525. Epub 2021 Sep 13. Aging (Albany NY). 2021. PMID: 34517343 Free PMC article.
-
Cuproplasia and cuproptosis, two sides of the coin.Cancer Commun (Lond). 2025 May;45(5):505-524. doi: 10.1002/cac2.70001. Epub 2025 Jan 25. Cancer Commun (Lond). 2025. PMID: 39865459 Free PMC article. Review.
References
-
- Madeo F, Eisenberg T, Pietrocola F, Kroemer G. Spermidine in health and disease. Science. 2018;359:pii: eaan2788. - PubMed
-
- Eisenberg T, Knauer H, Schauer A, Buttner S, Ruckenstuhl C, Carmona-Gutierrez D, et al. Induction of autophagy by spermidine promotes longevity. Nat Cell Biol. 2009;11:1305–14. - PubMed
-
- Kiechl S, Pechlaner R, Willeit P, Notdurfter M, Paulweber B, Willeit K, et al. Higher spermidine intake is linked to lower mortality: a prospective population-based study. Am J Clin Nutr. 2018;108:371–80.. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
