

Human serum proteins bind to Sporothrix schenckii conidia with differential effects on phagocytosis
- PMID: 32382937
- PMCID: PMC7966614
- DOI: 10.1007/s42770-020-00276-3
Human serum proteins bind to Sporothrix schenckii conidia with differential effects on phagocytosis
Abstract
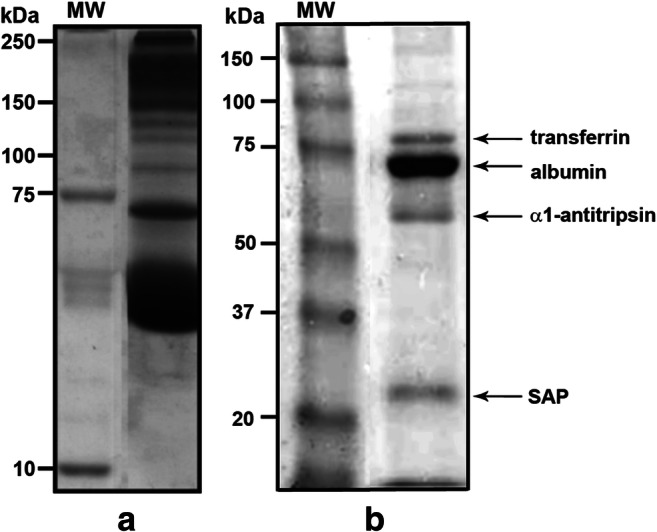
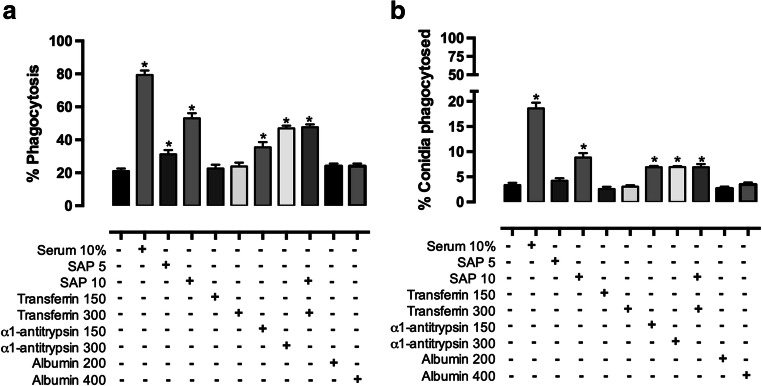
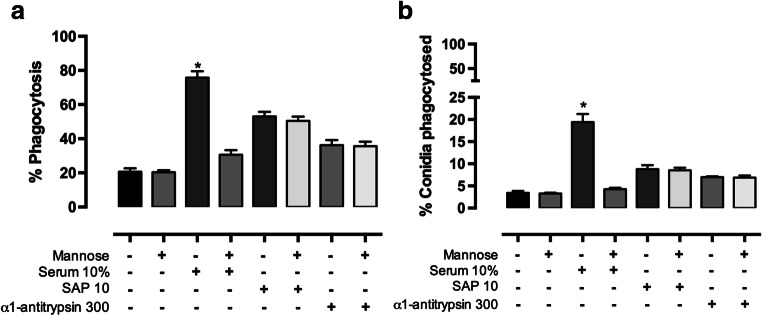
Serum is an important source of proteins that interact with pathogens. Once bound to the cell surface, serum proteins can stimulate the innate immune system. The phagocytosis of Sporothrix schenckii conidia by human macrophages is activated through human serum opsonisation. In this study, we have attempted to characterise human blood serum proteins that bind to the cell wall of S. schenckii conidia. We systematically observed the same four proteins independent of the plasma donor: albumin, serum amyloid protein (SAP), α-1 antitrypsin (AAT), and transferrin were identified with the help of tandem mass spectrometry. Phagocytosis depended on the concentration of the SAP or α-1 antitrypsin that was used to opsonise the conidia; however, transferrin or albumin did not have any effect on conidia internalisation. The presence of mannose did not affect macrophage phagocytosis of the conidia opsonised with SAP or α-1 antitrypsin, which suggests that these proteins are not recognised by the mannose receptor.
Keywords: Conidia; Macrophage; Phagocytosis; Serum amyloid protein; Sporotrichosis; α-1 antitrypsin.
Figures
References
-
- Zhang Y, Hagen F, Stielow B, Rodrigues AM, Samerpitak K, Zhou X, Feng P, Yang L, Chen M, Deng S, Li S, Liao W, Li R, Li F, Meis JF, Guarro J, Teixeira M, al-Zahrani HS, de Camargo ZP, Zhang L, de Hoog GS. Phylogeography and evolutionary patterns in Sporothrix spanning more than 14 000 human and animal case reports. Persoonia. 2015;35:1–20. doi: 10.3767/003158515x687416. - DOI - PMC - PubMed
-
- Morgado FN, Schubach AO, Pimentel MI, Lyra MR, Vasconcellos ÉCF, Valete-Rosalino CM, Conceição-Silva F. Is there any difference between the in situ and systemic IL-10 and IFN-γ production when clinical forms of cutaneous sporotrichosis are compared? PLoS One. 2016;11:e0162764. doi: 10.1371/journal.pone.0162764. - DOI - PMC - PubMed
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
