

Budding yeast complete DNA synthesis after chromosome segregation begins
- PMID: 32385287
- PMCID: PMC7210879
- DOI: 10.1038/s41467-020-16100-3
Budding yeast complete DNA synthesis after chromosome segregation begins
Abstract
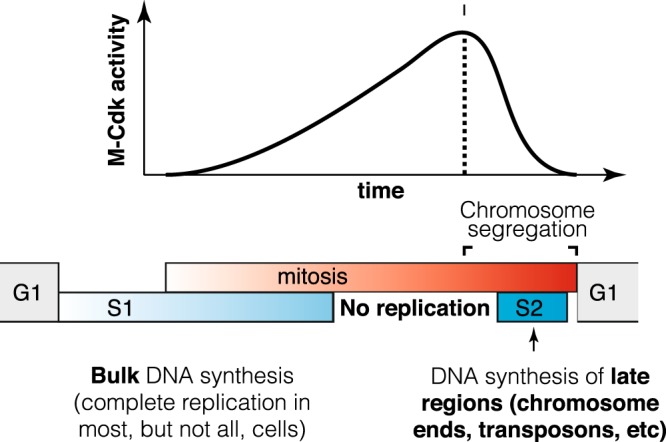
To faithfully transmit genetic information, cells must replicate their entire genome before division. This is thought to be ensured by the temporal separation of replication and chromosome segregation. Here we show that in 20-40% of unperturbed yeast cells, DNA synthesis continues during anaphase, late in mitosis. High cyclin-Cdk activity inhibits DNA synthesis in metaphase, and the decrease in cyclin-Cdk activity during mitotic exit allows DNA synthesis to finish at subtelomeric and some difficult-to-replicate regions. DNA synthesis during late mitosis correlates with elevated mutation rates at subtelomeric regions, including copy number variation. Thus, yeast cells temporally overlap DNA synthesis and chromosome segregation during normal growth, possibly allowing cells to maximize population-level growth rate while simultaneously exploring greater genetic space.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Morgan, D. O. The Cell Cycle: Principles of Control (Primers in Biology) (Sinauer Associates, Inc., 2006).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
