

Cell-specific metabolomic responses to injury: novel insights into blood-brain barrier modulation
- PMID: 32385409
- PMCID: PMC7210983
- DOI: 10.1038/s41598-020-64722-w
Cell-specific metabolomic responses to injury: novel insights into blood-brain barrier modulation
Abstract
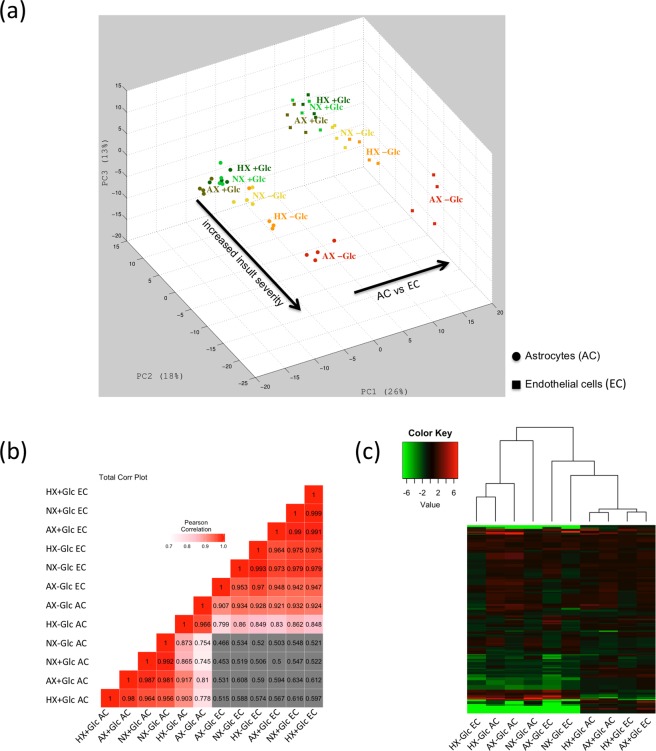
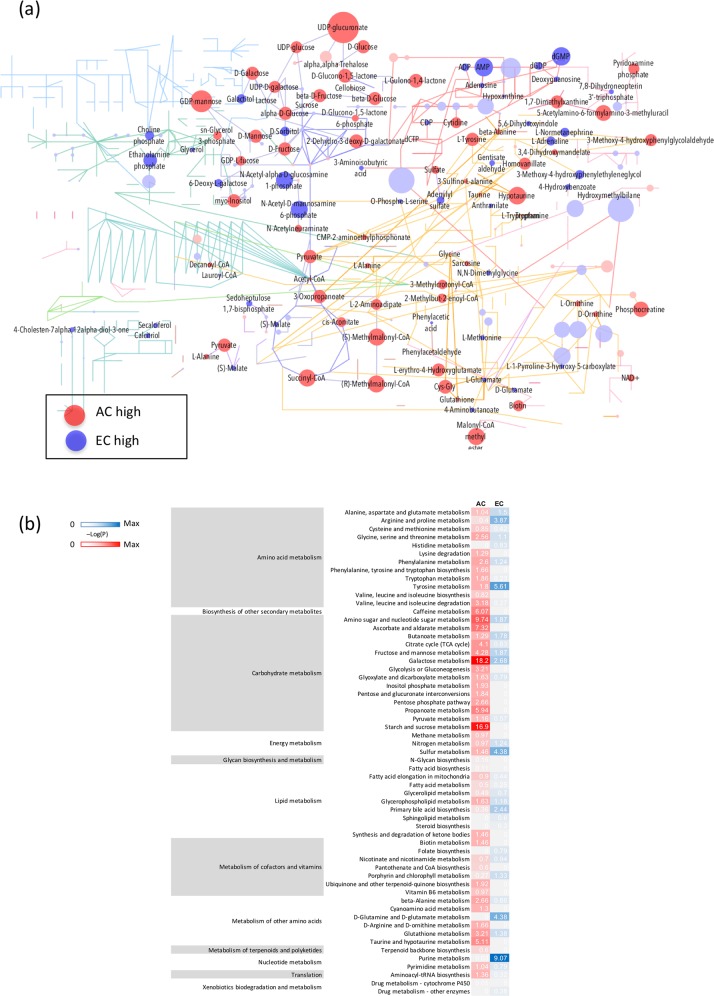
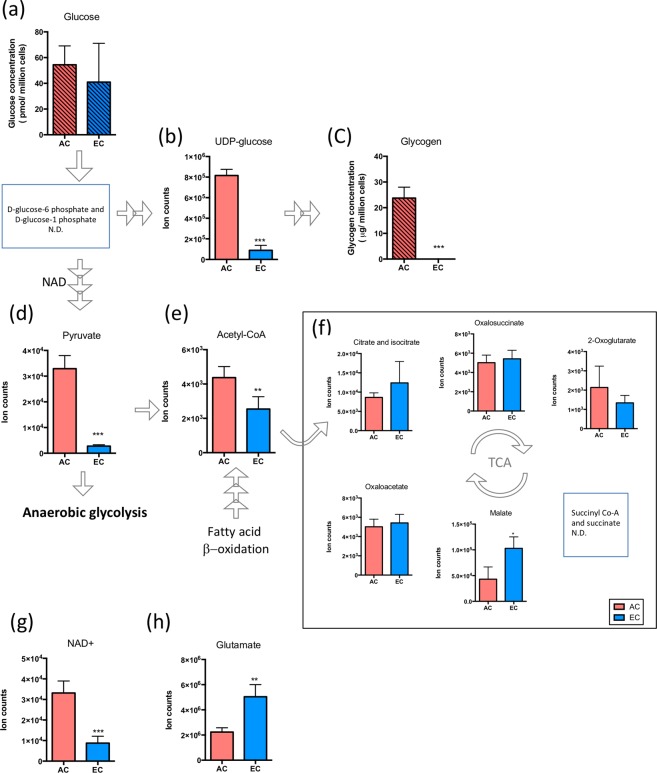
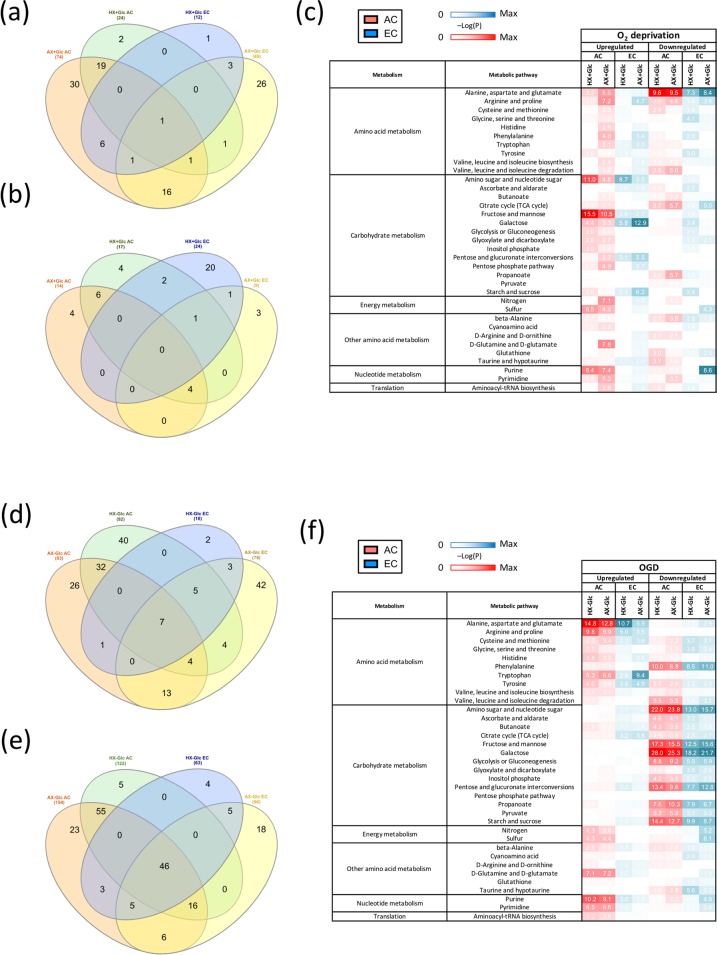
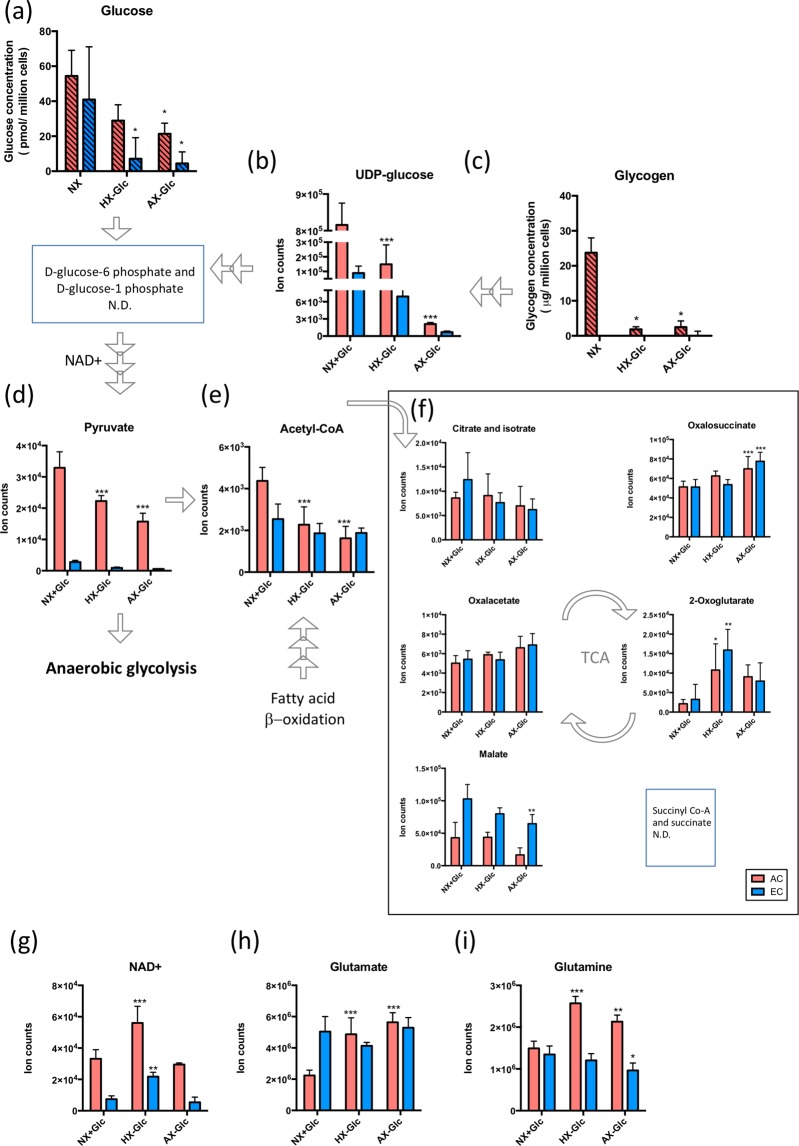
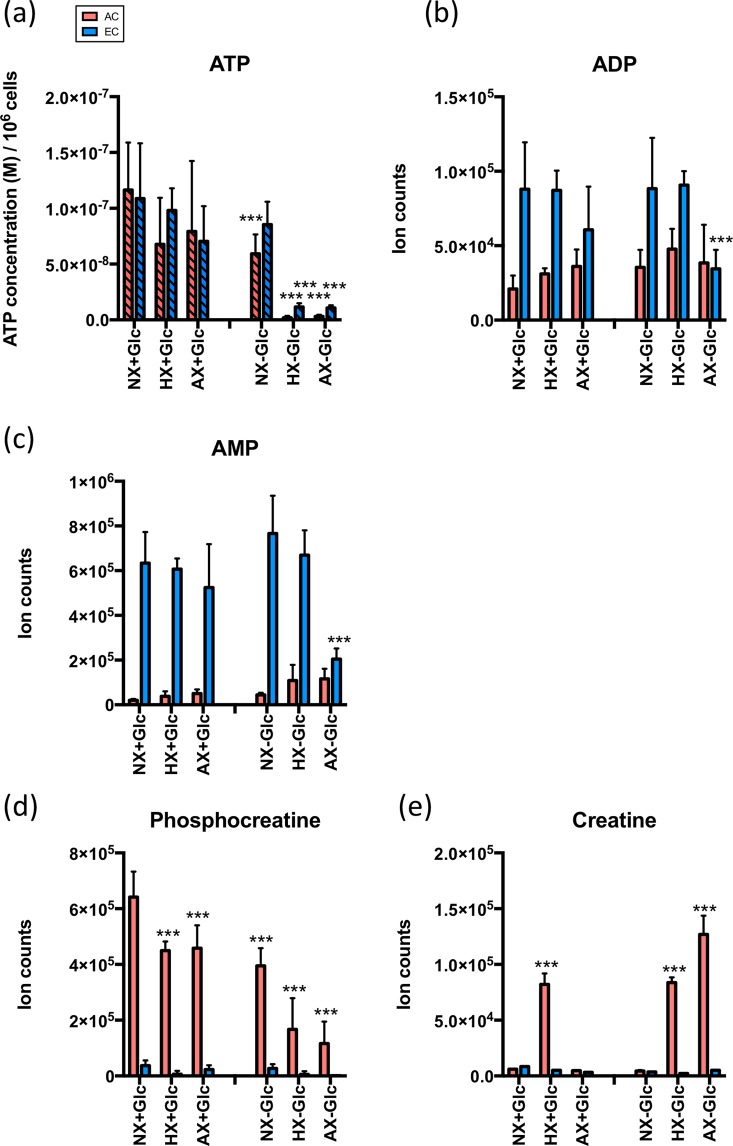
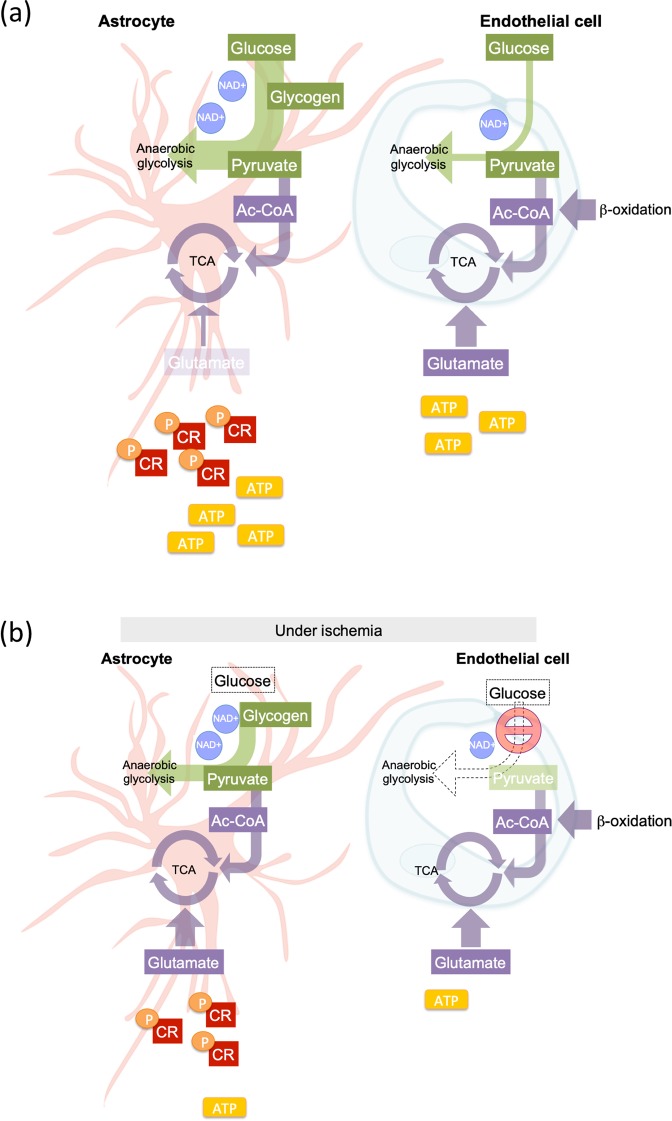
On one hand blood-brain barrier (BBB) disturbance aggravates disease progression, on the other it prevents drug access and impedes therapeutic efficacy. Effective ways to modulate barrier function and resolve these issues are sorely needed. Convinced that better understanding of cell-oriented BBB responses could provide valuable insight, and the fact that metabolic dysregulation is prominent in many vascular-related pathological processes associated with BBB disturbance, we hypothesized that differential cell-specific metabolic adaptation majorly influences physiological and pathological barrier functionality. Untargeted liquid chromatography-mass spectrometry (LC-MS) metabolomic profiling was used to obtain individual biochemical fingerprints of primary astrocytes (AC) and brain endothelial cells (EC) during normoxic conditions and increasing hypoxic/ischemic injury and thus a functional readout of cell status. Bioinformatic analyses showed each cell had a distinct metabolic signature. Corroborating their roles in BBB and CNS protection, AC showed an innate ability to dynamically alter their metabolome depending on the insult. Surprisingly, in complete contrast, EC largely maintained their normoxic characteristics in injury situations and their profiles diverged from those of non-brain origin. Tissue specificity/origin is clearly important when considering EC responses. Focusing on energy capacity and utilization we discuss how cell-specific metabolic adaptive capabilities could influence vascular stability and the possibility that altering metabolite levels may be an effective way to modulate brain EC function. Overall this work novel insight into cell-associated metabolic changes, and provides a powerful resource for understanding BBB changes during different injury scenarios.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
