

IL-7 induces sCD127 release and mCD127 downregulation in human CD8+ T cells by distinct yet overlapping mechanisms, both of which are impaired in HIV infection
- PMID: 32390135
- PMCID: PMC7586945
- DOI: 10.1002/eji.201948453
IL-7 induces sCD127 release and mCD127 downregulation in human CD8+ T cells by distinct yet overlapping mechanisms, both of which are impaired in HIV infection
Abstract
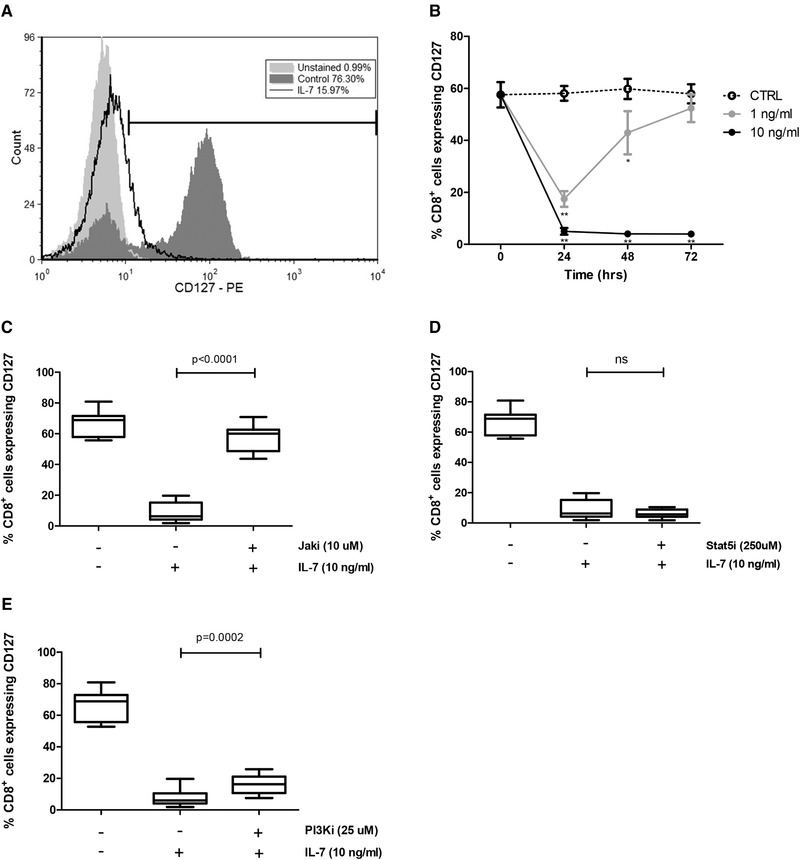
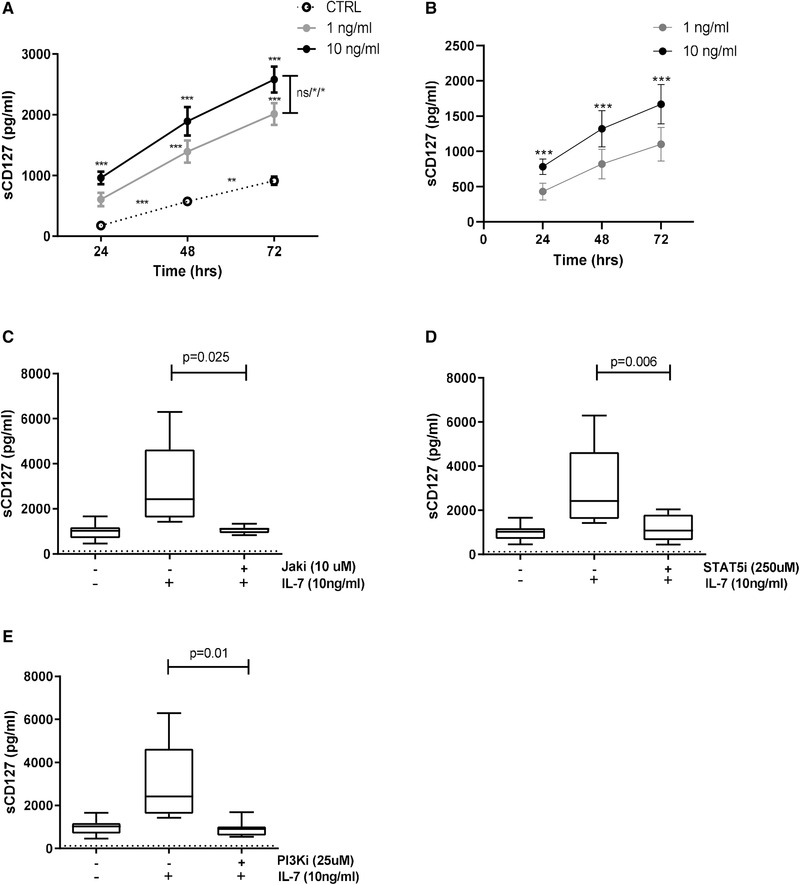
The IL-7 receptor specific α chain, CD127, can be expressed both as a membrane-associated (mCD127) and a soluble form (sCD127), however, the mechanisms involved in their regulation remain to be defined. We first demonstrated in primary human CD8+ T cells that IL-7-induced downregulation of mCD127 expression is dependent on JAK and PI3K signaling, whereas IL-7-induced sCD127 release is also mediated by STAT5. Following stimulation with IL-7, expression of alternatively spliced variants of the CD127 gene, sCD127 mRNA, is reduced, but to a lesser degree than the full-length gene. Evaluation of the role of proteases revealed that MMP-9 was involved in sCD127 release, without affecting the expression of mCD127, suggesting it does not induce direct shedding from the cell surface. Since defects in the IL-7/CD127 pathway occur in various diseases, including HIV, we evaluated CD8+ T cells derived from HAART-treated HIV-infected individuals and found that IL-7-induced (1) downregulation of mCD127, (2) release of sCD127, and (3) expression of the sCD127 mRNA were all impaired. Expression of mCD127 and sCD127 is, therefore, regulated by distinct, but overlapping, mechanisms and their impairment in HIV infection contributes to our understanding of the CD8+ T cell dysfunction that persists despite effective antiretroviral therapy.
Keywords: CD127; CD8+ T cells; HAART; HIV; IL-7.
© 2020 The Authors. European Journal of Immunology published by WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Conflict of interest statement
The authors declare no commercial or financial conflict of interest.
Figures



References
-
- Pallard C., Stegmann A. P., van Kleffens T., Smart F., Venkitaraman A. and Spits H., Distinct roles of the phosphatidylinositol 3‐kinase and STAT5 pathways in IL‐7‐mediated development of human thymocyte precursors. Immunity 1999. 10: 525–535. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
