

Protamine loops DNA in multiple steps
- PMID: 32392345
- PMCID: PMC7293030
- DOI: 10.1093/nar/gkaa365
Protamine loops DNA in multiple steps
Abstract
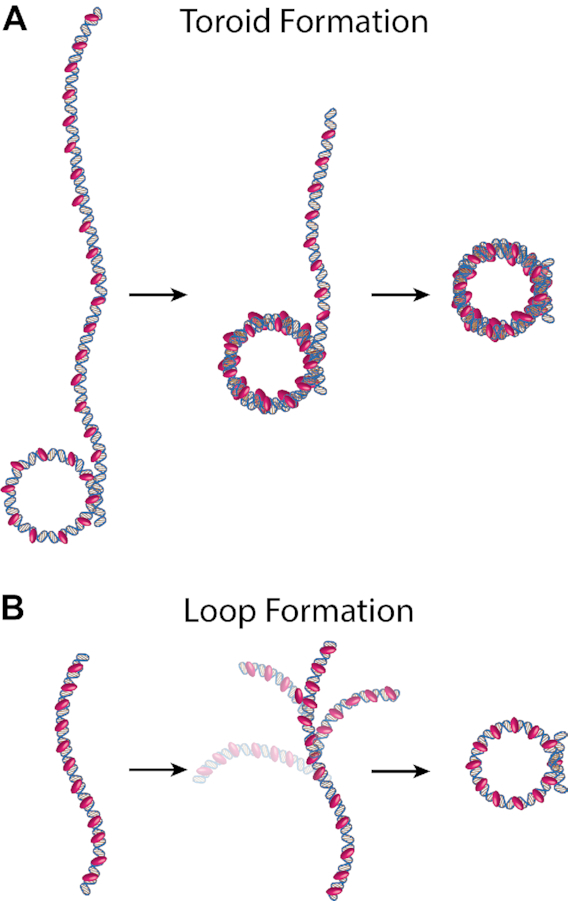
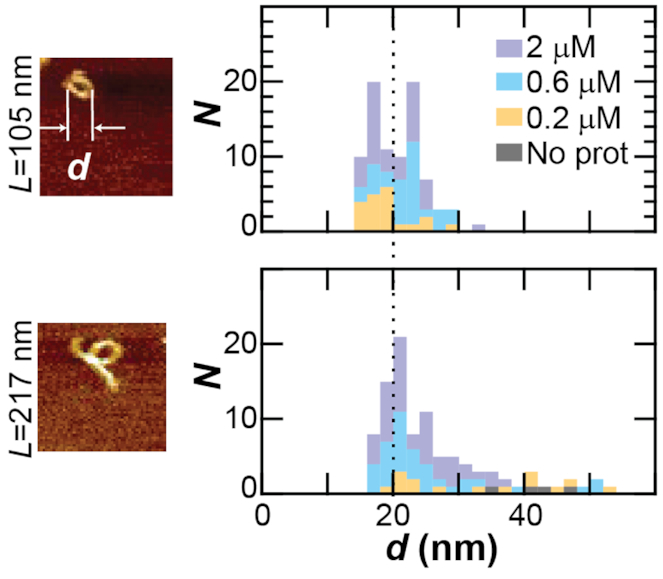
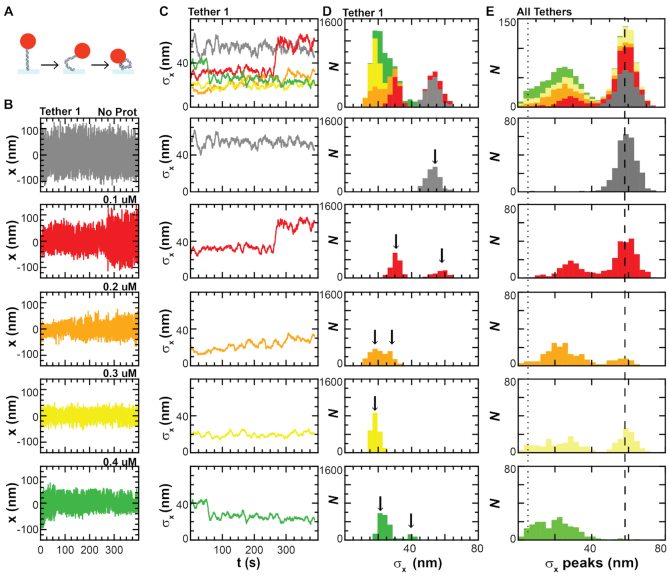
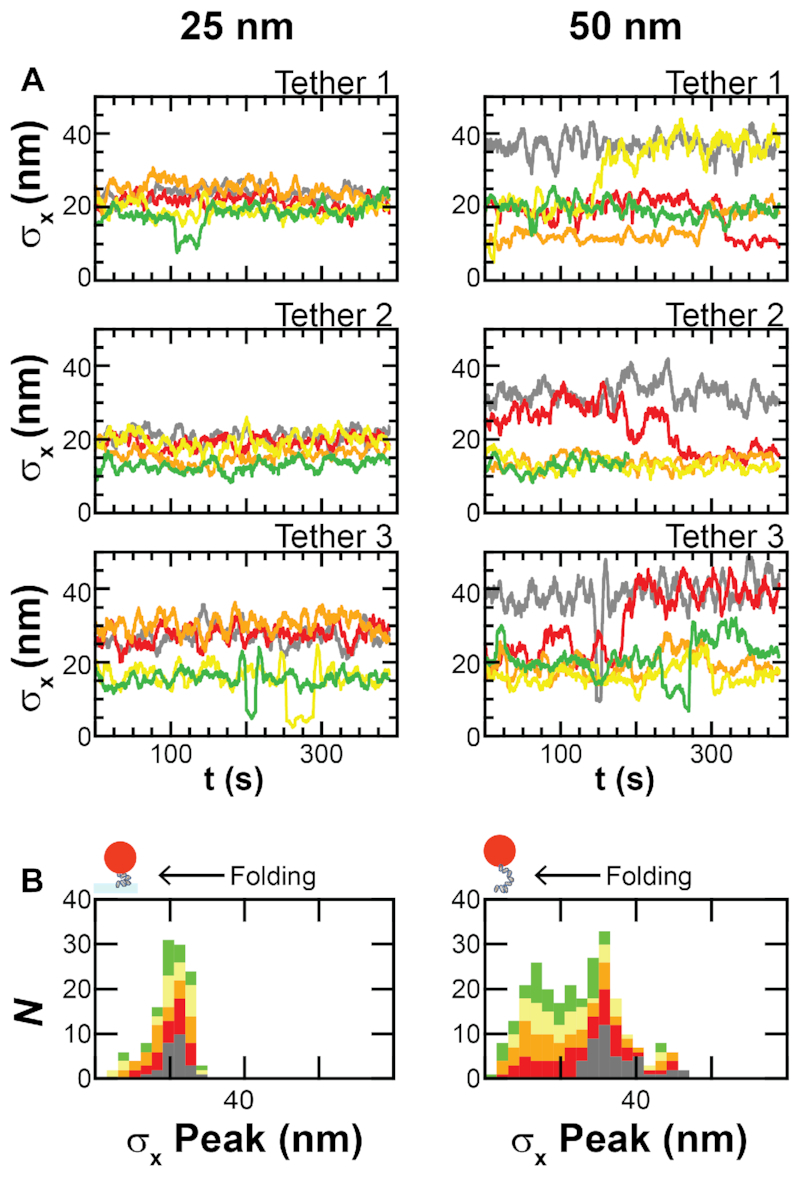
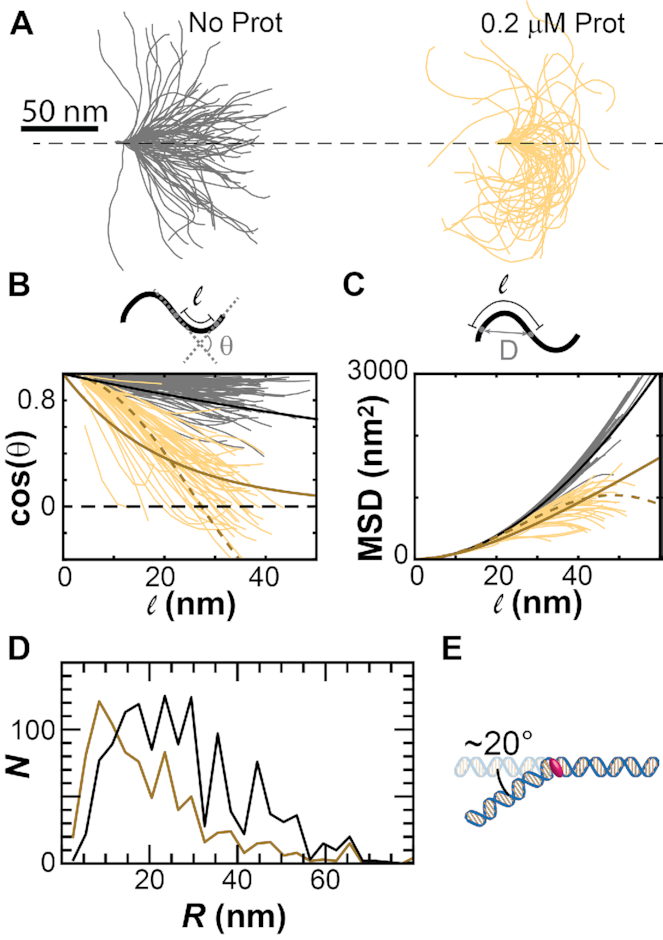
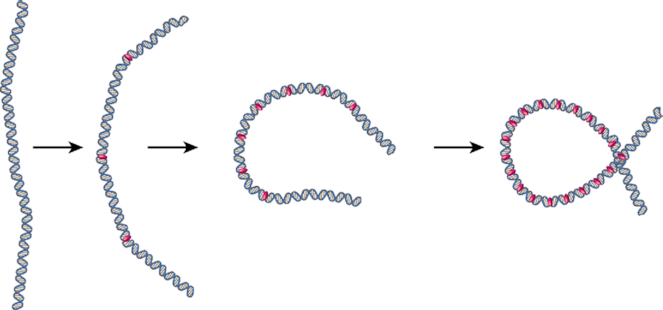
Protamine proteins dramatically condense DNA in sperm to almost crystalline packing levels. Here, we measure the first step in the in vitro pathway, the folding of DNA into a single loop. Current models for DNA loop formation are one-step, all-or-nothing models with a looped state and an unlooped state. However, when we use a Tethered Particle Motion (TPM) assay to measure the dynamic, real-time looping of DNA by protamine, we observe the presence of multiple folded states that are long-lived (∼100 s) and reversible. In addition, we measure folding on DNA molecules that are too short to form loops. This suggests that protamine is using a multi-step process to loop the DNA rather than a one-step process. To visualize the DNA structures, we used an Atomic Force Microscopy (AFM) assay. We see that some folded DNA molecules are loops with a ∼10-nm radius and some of the folded molecules are partial loops-c-shapes or s-shapes-that have a radius of curvature of ∼10 nm. Further analysis of these structures suggest that protamine is bending the DNA to achieve this curvature rather than increasing the flexibility of the DNA. We therefore conclude that protamine loops DNA in multiple steps, bending it into a loop.
© The Author(s) 2020. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures

References
-
- Pogany G.C., Corzett M., Weston S., Balhorn R.. DNA and protein content of mouse sperm. implications regarding sperm chromatin structure. Exp. Cell Res. 1981; 136:127–136. - PubMed
-
- Hud N.V., Milanovich F.P., Balhorn R.. Evidence of novel secondary structure in DNA-bound protamine is revealed by Raman spectroscopy. Biochemistry. 1994; 33:7528–7535. - PubMed
-
- Teif V.B., Bohinc K.. Condensed DNA: condensing the concepts. Prog. Biophys. Mol. Biol. 2011; 105:208–222. - PubMed
-
- Sotolongo B., Lino E., Ward W.S.. Ability of hamster spermatozoa to digest their own DNA. Biol. Reprod. 2003; 69:2029–2035. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
