

Hippocampal Network Reorganization Underlies the Formation of a Temporal Association Memory
- PMID: 32392472
- PMCID: PMC7643350
- DOI: 10.1016/j.neuron.2020.04.013
Hippocampal Network Reorganization Underlies the Formation of a Temporal Association Memory
Abstract
Episodic memory requires linking events in time, a function dependent on the hippocampus. In "trace" fear conditioning, animals learn to associate a neutral cue with an aversive stimulus despite their separation in time by a delay period on the order of tens of seconds. But how this temporal association forms remains unclear. Here we use two-photon calcium imaging of neural population dynamics throughout the course of learning and show that, in contrast to previous theories, hippocampal CA1 does not generate persistent activity to bridge the delay. Instead, learning is concomitant with broad changes in the active neural population. Although neural responses were stochastic in time, cue identity could be read out from population activity over longer timescales after learning. These results question the ubiquity of seconds-long neural sequences during temporal association learning and suggest that trace fear conditioning relies on mechanisms that differ from persistent activity accounts of working memory.
Keywords: calcium imaging; hippocampus; learning; memory; population coding; trace fear conditioning.
Copyright © 2020. Published by Elsevier Inc.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
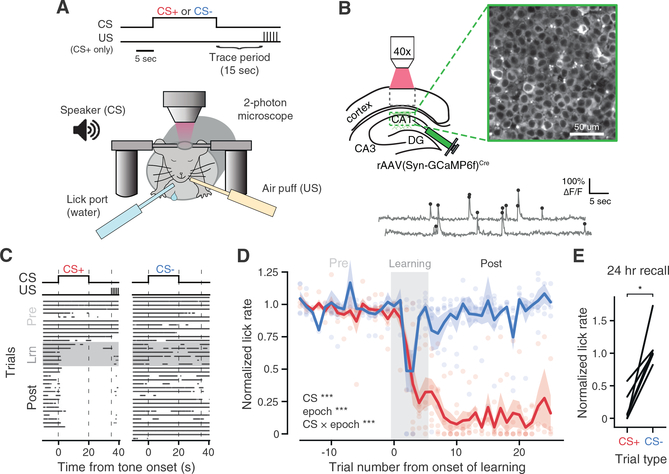
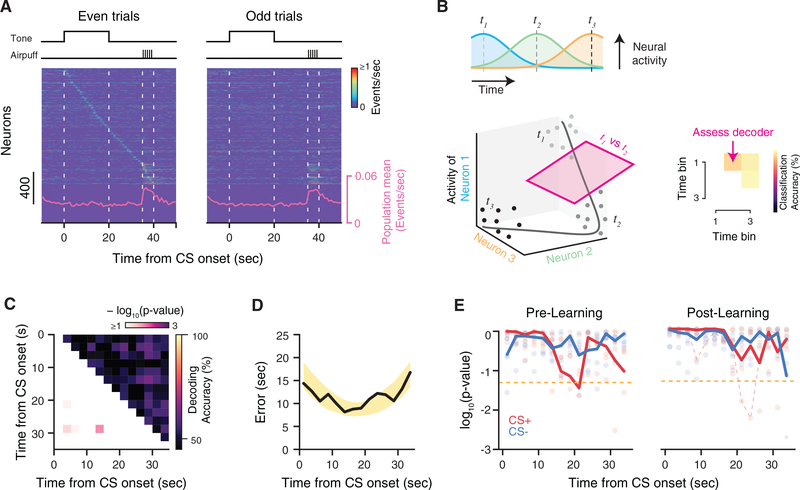
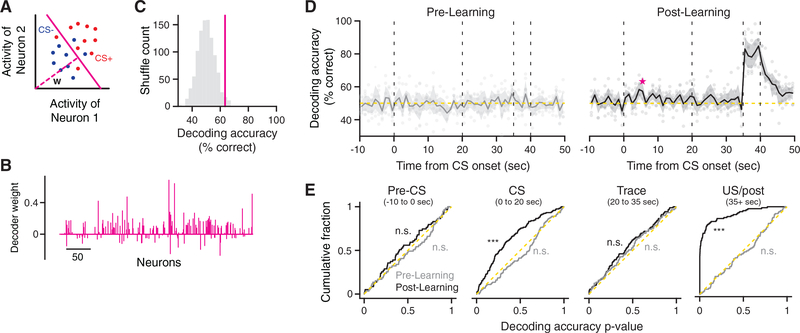
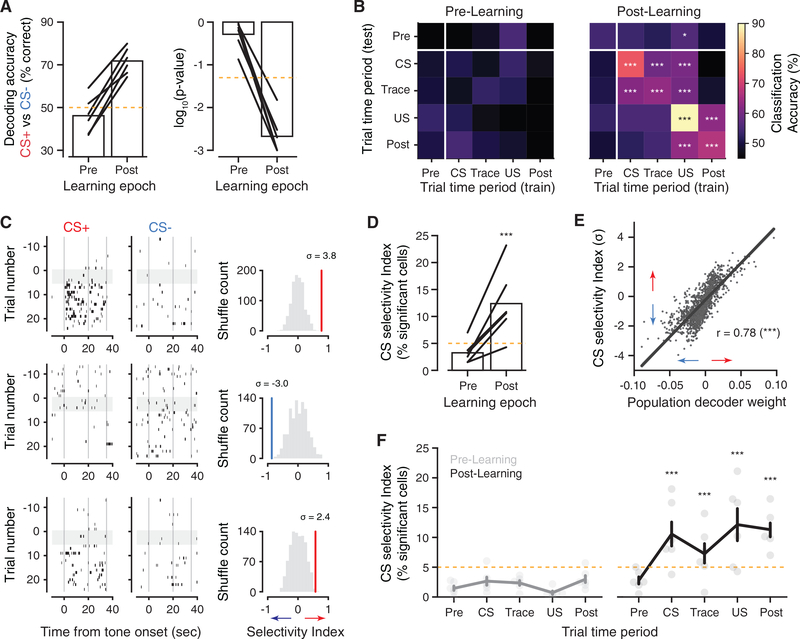
Comment in
-
Tracing a Path for Memory in the Hippocampus.Neuron. 2020 Jul 22;107(2):199-201. doi: 10.1016/j.neuron.2020.06.034. Neuron. 2020. PMID: 32702341 Free PMC article.
References
-
- Amit DJ, and Brunel N (1997). Model of global spontaneous activity and local structured activity during delay periods in the cerebral cortex. Cereb. Cortex 7, 237–252. - PubMed
-
- Barak O, and Tsodyks M (2014). Working models of working memory. Curr. Opin. Neurobiol. 25, 20–24. - PubMed
-
- Benna MK, and Fusi S (2016). Computational principles of synaptic memory consolidation. Nat. Neurosci. 19, 1697–1706. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
