

AURKA destruction is decoupled from its activity at mitotic exit but is essential to suppress interphase activity
- PMID: 32393600
- PMCID: PMC7328152
- DOI: 10.1242/jcs.243071
AURKA destruction is decoupled from its activity at mitotic exit but is essential to suppress interphase activity
Abstract
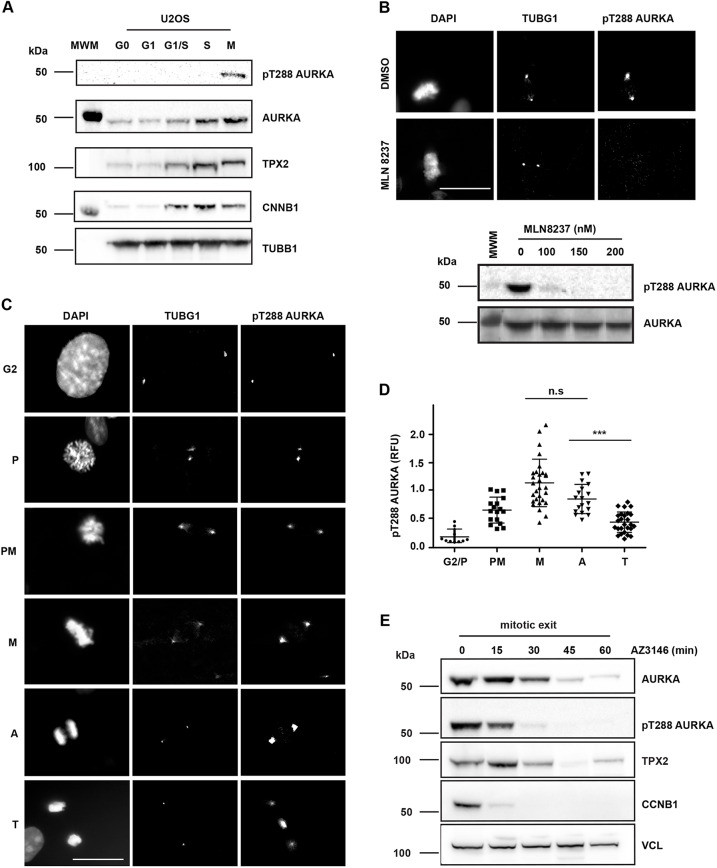
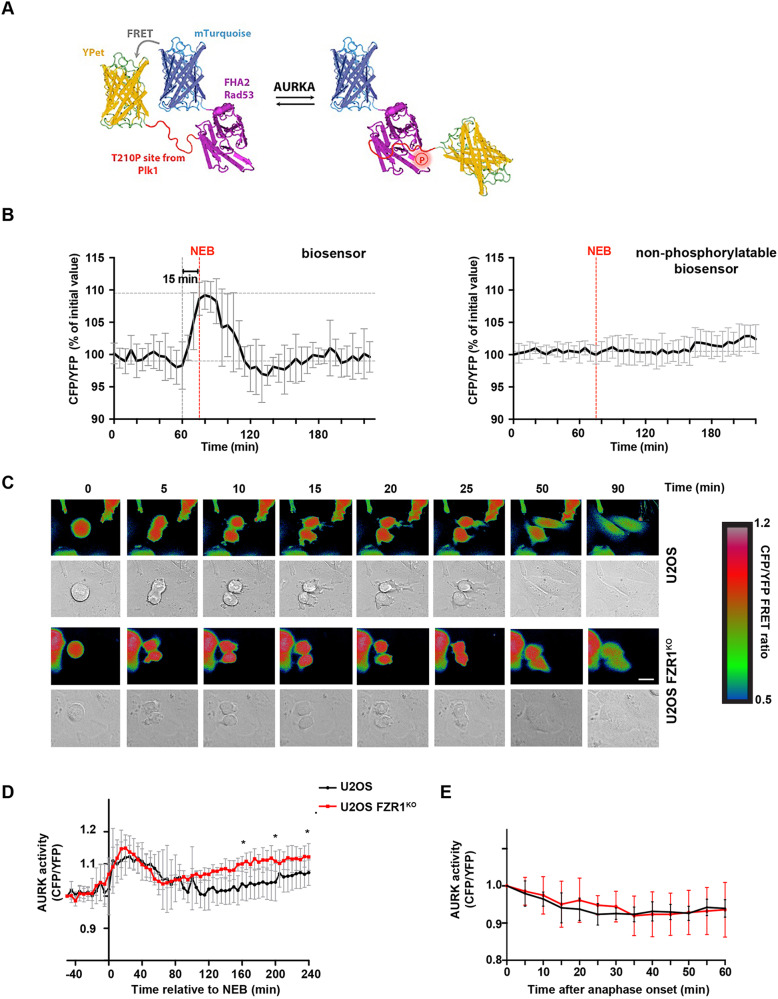
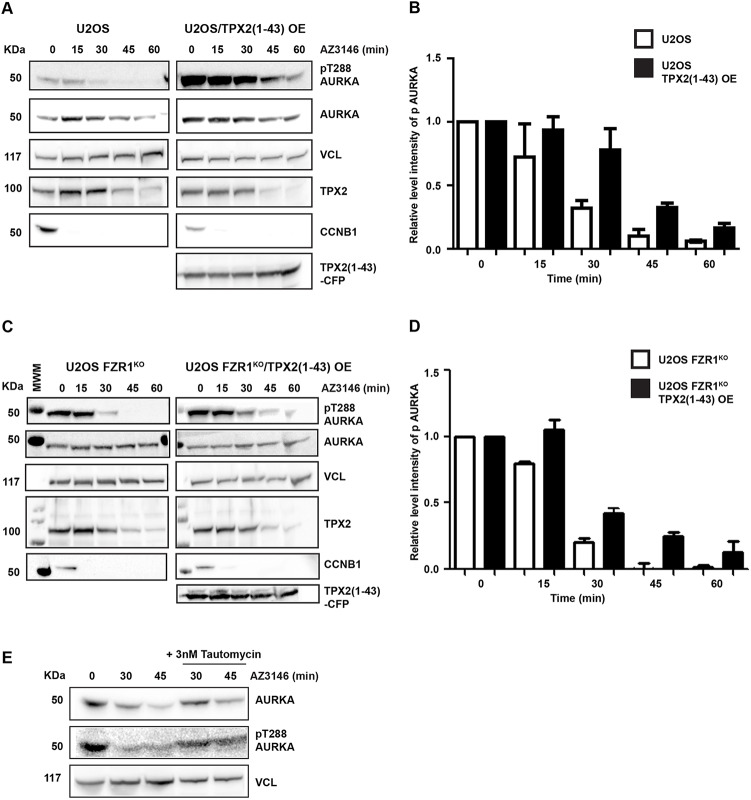
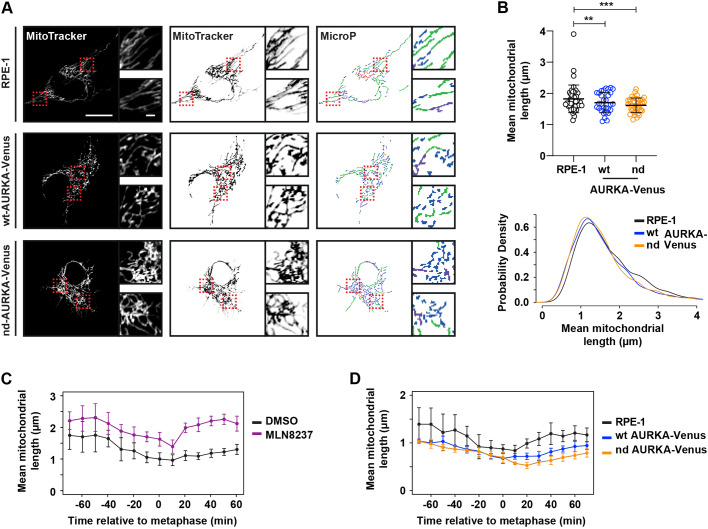
Activity of AURKA is controlled through multiple mechanisms including phosphorylation, ubiquitin-mediated degradation and allosteric interaction with TPX2. Activity peaks at mitosis, before AURKA is degraded during and after mitotic exit in a process strictly dependent on the APC/C coactivator FZR1. We used FZR1 knockout cells (FZR1KO) and a novel FRET-based AURKA biosensor to investigate how AURKA activity is regulated in the absence of destruction. We found that AURKA activity in FZR1KO cells dropped at mitotic exit as rapidly as in parental cells, despite absence of AURKA destruction. Unexpectedly, TPX2 was degraded normally in FZR1KO cells. Overexpression of an N-terminal TPX2 fragment sufficient for AURKA binding, but that is not degraded at mitotic exit, caused delay in AURKA inactivation. We conclude that inactivation of AURKA at mitotic exit is determined not by AURKA degradation but by degradation of TPX2 and therefore is dependent on CDC20 rather than FZR1. The biosensor revealed that FZR1 instead suppresses AURKA activity in interphase and is critically required for assembly of the interphase mitochondrial network after mitosis.This article has an associated First Person interview with the first authors of the paper.
Keywords: Aurora A kinase; Cdh1; FZR1; Mitochondria; Mitosis; TPX2.
© 2020. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures


References
-
- Asteriti I. A., Di Cesare E., De Mattia F., Hilsenstein V., Neumann B., Cundari E., Lavia P. and Guarguaglini G. (2014). The Aurora-A inhibitor MLN8237 affects multiple mitotic processes and induces dose-dependent mitotic abnormalities and aneuploidy. Oncotarget 5, 6229-6242. 10.18632/oncotarget.2190 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
