

Biased signaling by endogenous opioid peptides
- PMID: 32393639
- PMCID: PMC7261131
- DOI: 10.1073/pnas.2000712117
Biased signaling by endogenous opioid peptides
Abstract
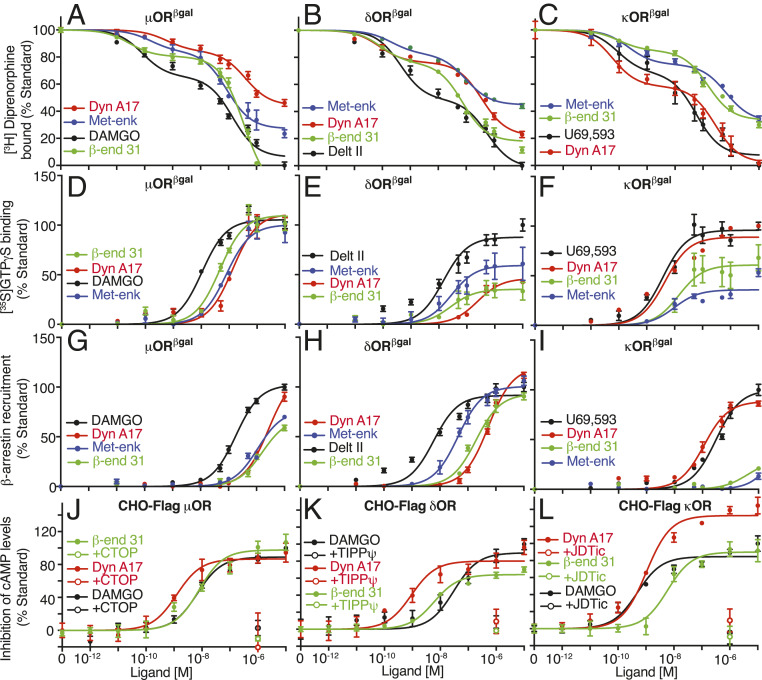
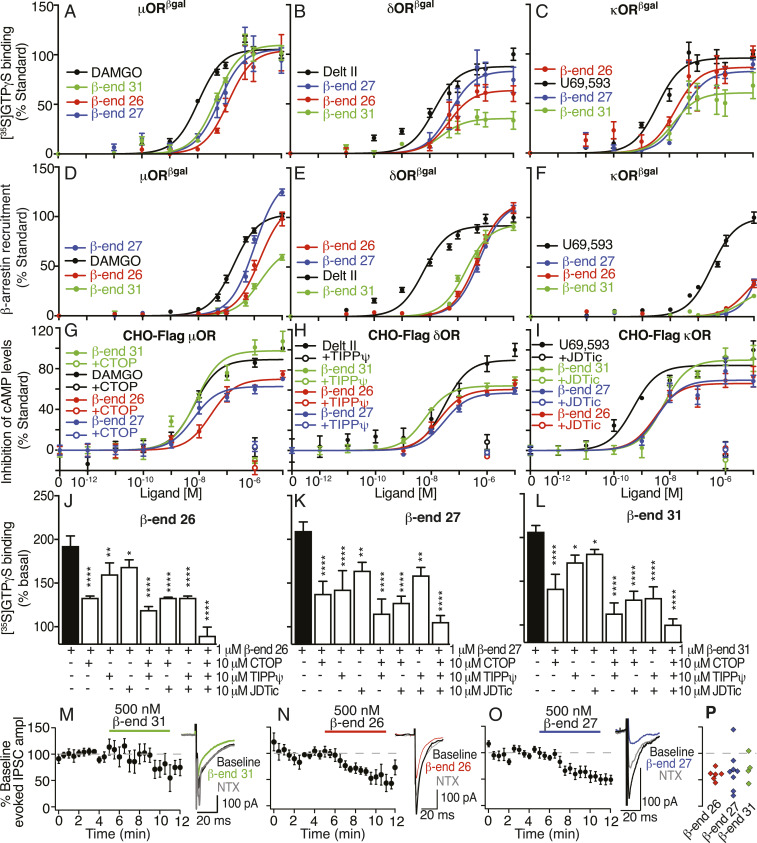
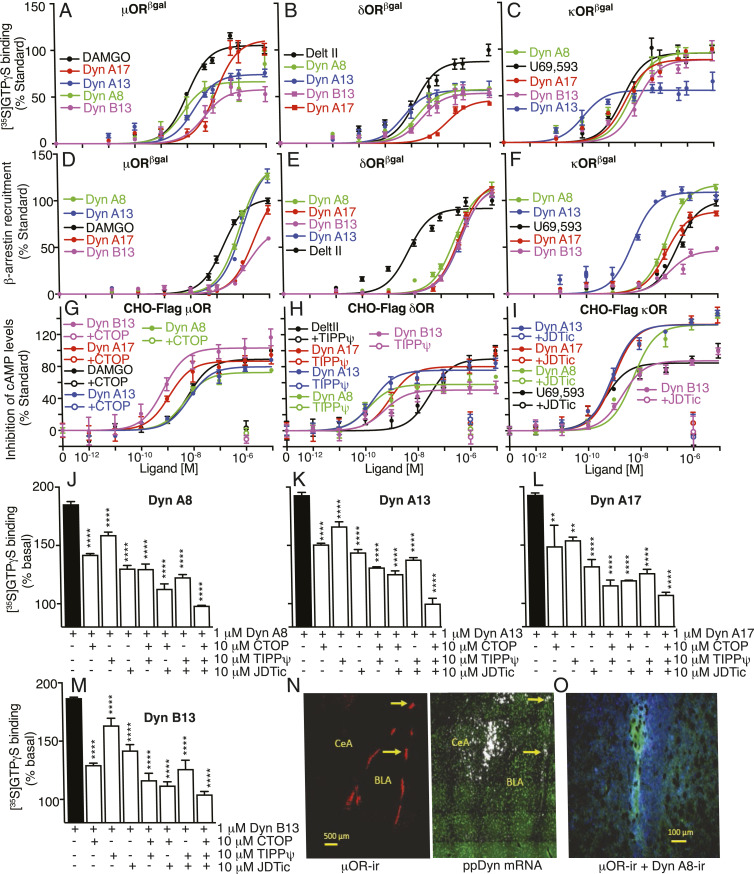
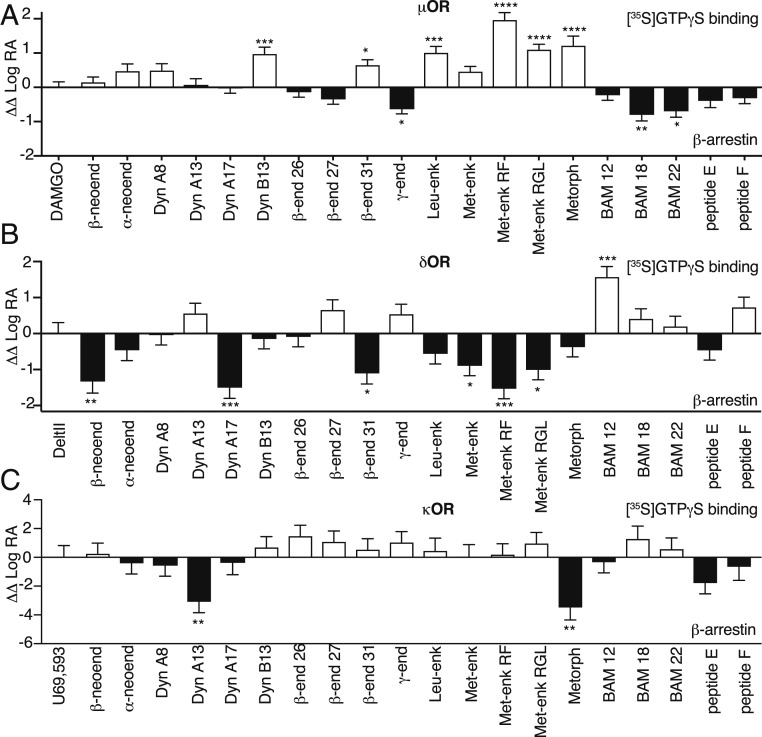
Opioids, such as morphine and fentanyl, are widely used for the treatment of severe pain; however, prolonged treatment with these drugs leads to the development of tolerance and can lead to opioid use disorder. The "Opioid Epidemic" has generated a drive for a deeper understanding of the fundamental signaling mechanisms of opioid receptors. It is generally thought that the three types of opioid receptors (μ, δ, κ) are activated by endogenous peptides derived from three different precursors: Proopiomelanocortin, proenkephalin, and prodynorphin. Posttranslational processing of these precursors generates >20 peptides with opioid receptor activity, leading to a long-standing question of the significance of this repertoire of peptides. Here, we address some aspects of this question using a technical tour de force approach to systematically evaluate ligand binding and signaling properties ([35S]GTPγS binding and β-arrestin recruitment) of 22 peptides at each of the three opioid receptors. We show that nearly all tested peptides are able to activate the three opioid receptors, and many of them exhibit agonist-directed receptor signaling (functional selectivity). Our data also challenge the dogma that shorter forms of β-endorphin do not exhibit receptor activity; we show that they exhibit robust signaling in cultured cells and in an acute brain slice preparation. Collectively, this information lays the groundwork for improved understanding of the endogenous opioid system that will help in developing more effective treatments for pain and addiction.
Keywords: GPCRs; biased agonism; opioid peptides; opioid receptors.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Stein C., Opioid receptors. Annu. Rev. Med. 67, 433–451 (2016). - PubMed
-
- Shenoy S. S., Lui F., Biochemistry, Endogenous Opioids (StatPearls, Treasure Island, FL, 2018). - PubMed
-
- Stein C., New concepts in opioid analgesia. Expert Opin. Investig. Drugs 27, 765–775 (2018). - PubMed
-
- Mansour A., Hoversten M. T., Taylor L. P., Watson S. J., Akil H., The cloned mu, delta and kappa receptors and their endogenous ligands: Evidence for two opioid peptide recognition cores. Brain Res. 700, 89–98 (1995). - PubMed
-
- Höllt V., Opioid peptide processing and receptor selectivity. Annu. Rev. Pharmacol. Toxicol. 26, 59–77 (1986). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
