

Stromal reactivity differentially drives tumour cell evolution and prostate cancer progression
- PMID: 32393869
- PMCID: PMC11000594
- DOI: 10.1038/s41559-020-1157-y
Stromal reactivity differentially drives tumour cell evolution and prostate cancer progression
Abstract
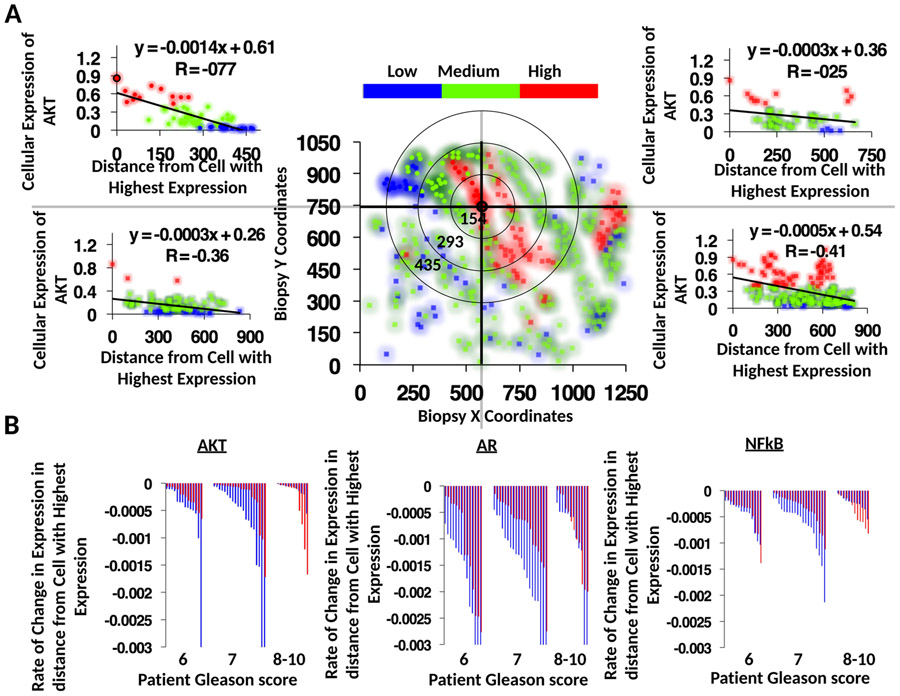
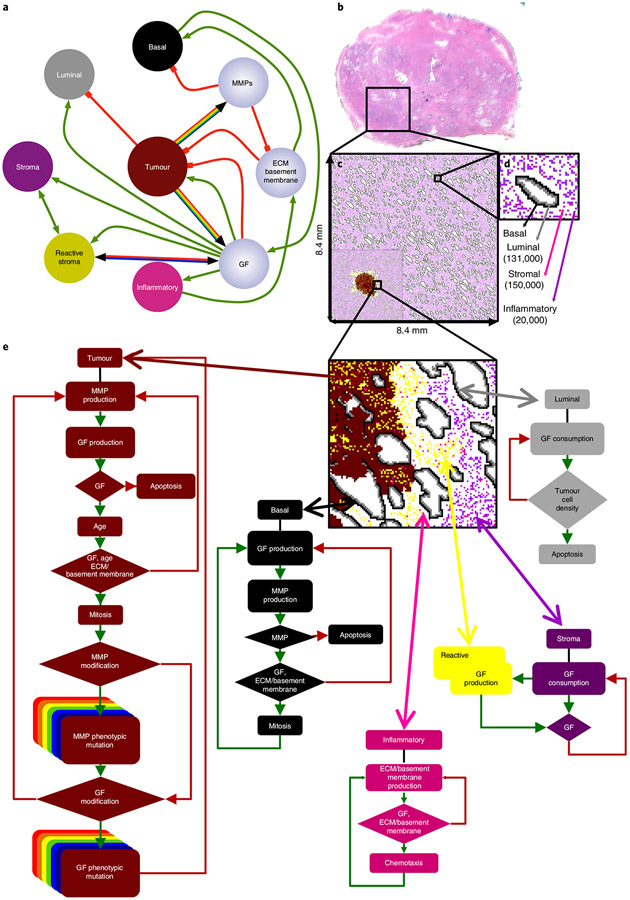
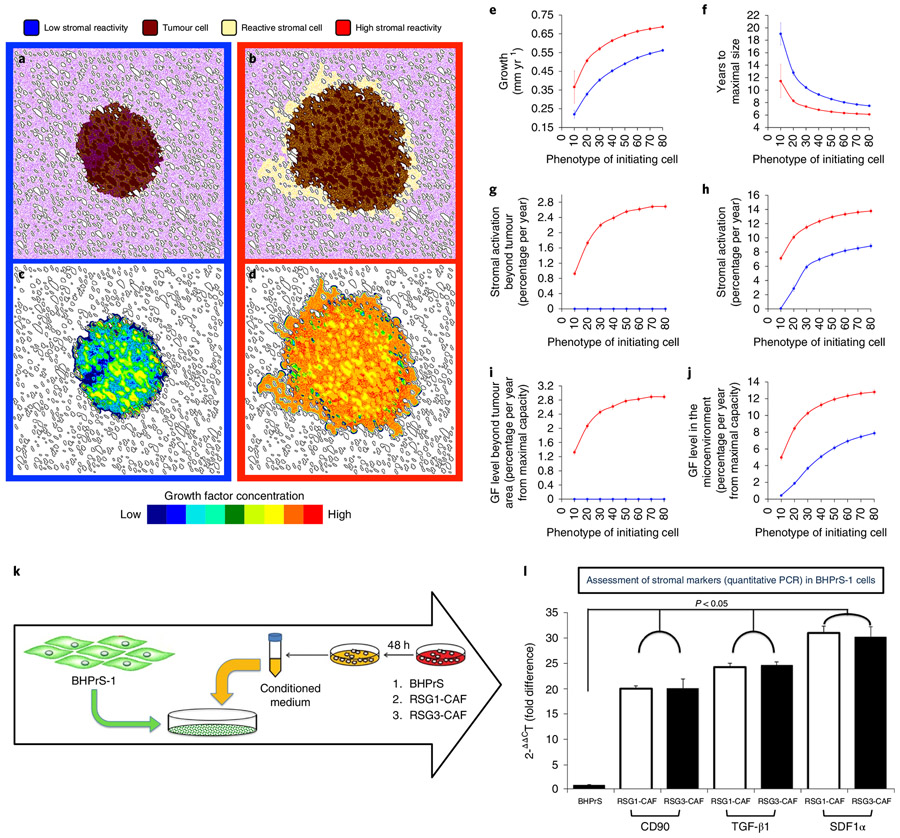
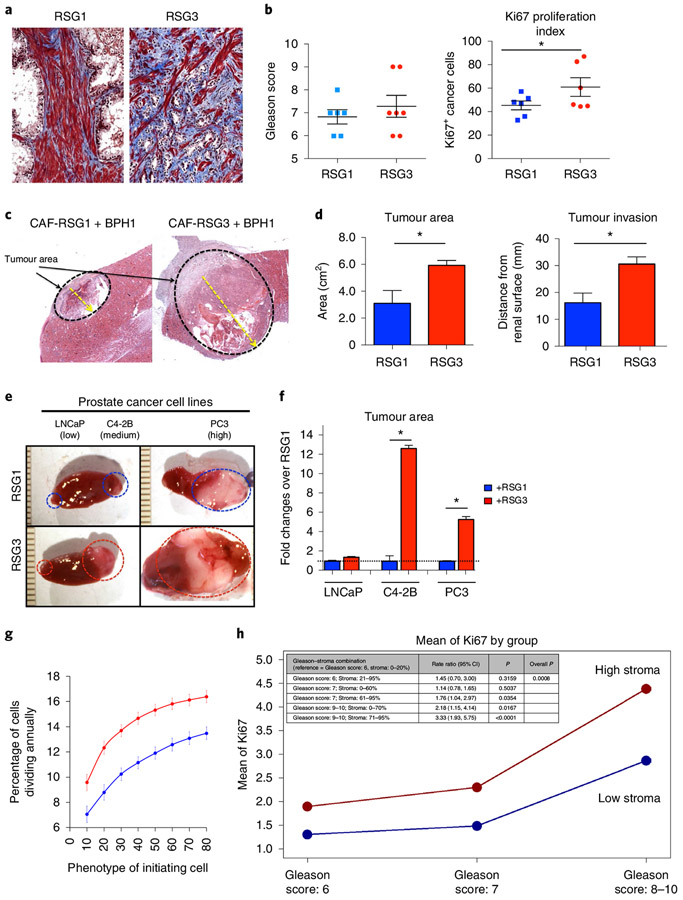
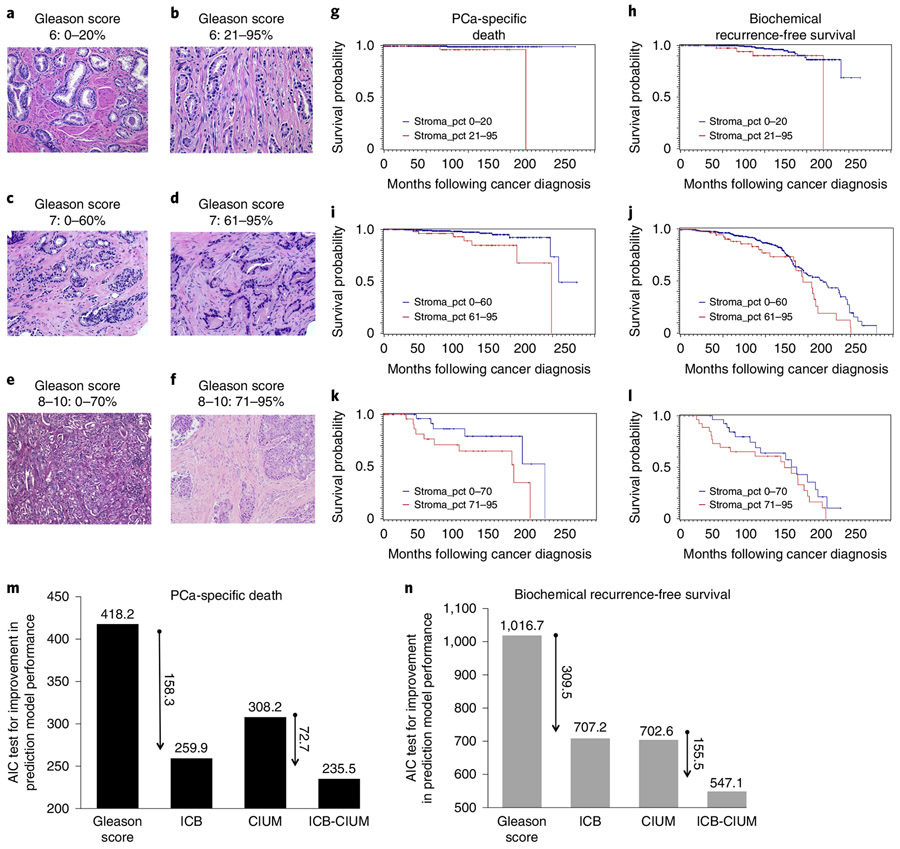
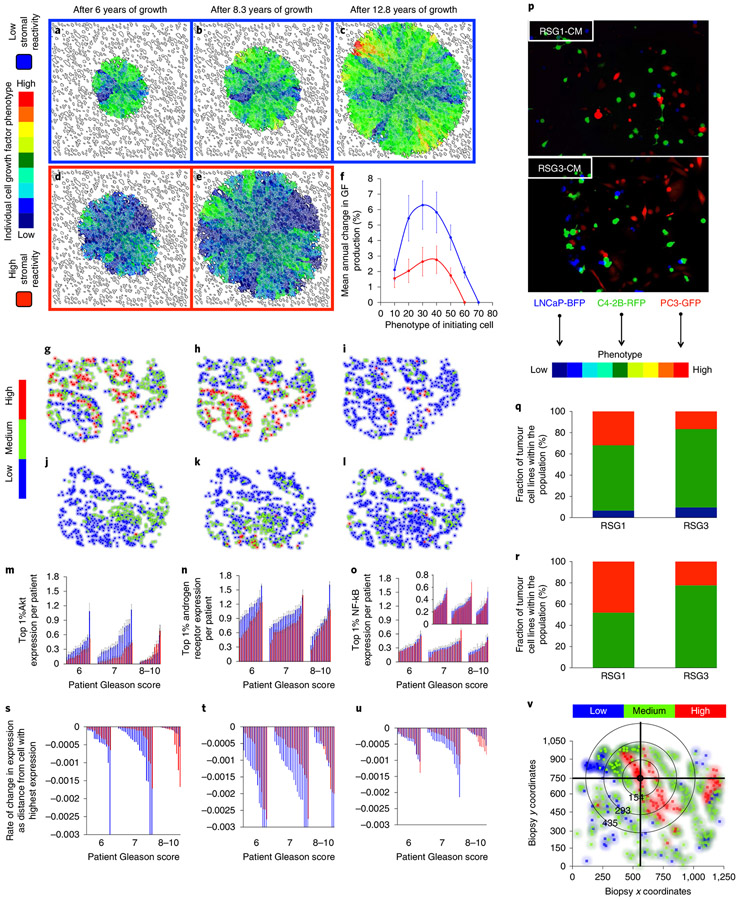
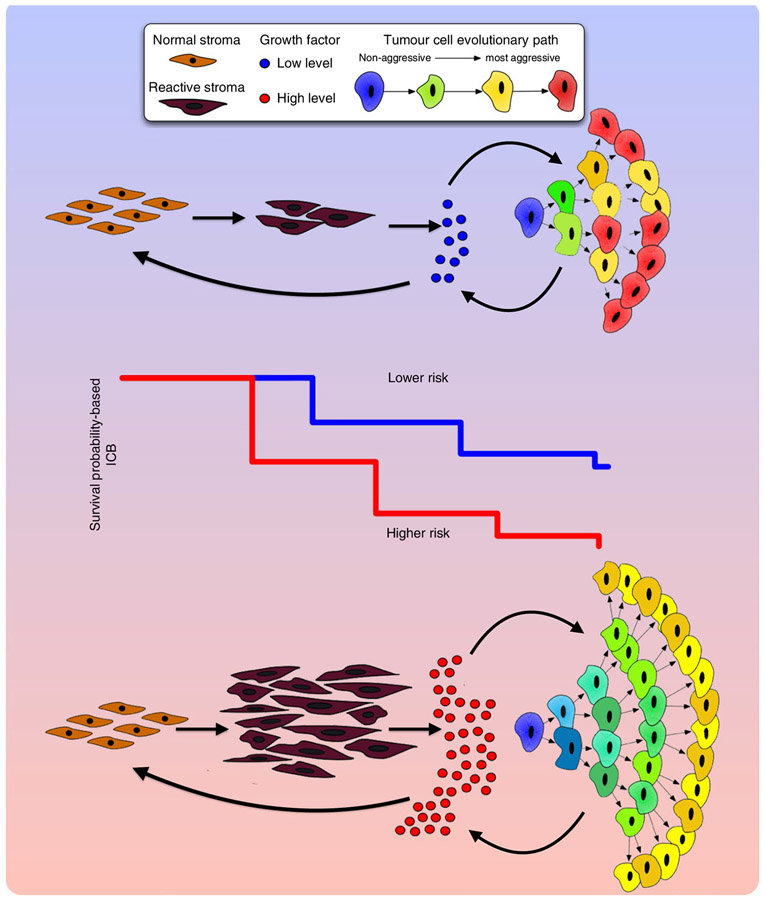
Prostate cancer (PCa) progression is a complex eco-evolutionary process driven by the feedback between evolving tumour cell phenotypes and microenvironmentally driven selection. To better understand this relationship, we used a multiscale mathematical model that integrates data from biology and pathology on the microenvironmental regulation of PCa cell behaviour. Our data indicate that the interactions between tumour cells and their environment shape the evolutionary dynamics of PCa cells and explain overall tumour aggressiveness. A key environmental determinant of this aggressiveness is the stromal ecology, which can be either inhibitory, highly reactive (supportive) or non-reactive (neutral). Our results show that stromal ecology correlates directly with tumour growth but inversely modulates tumour evolution. This suggests that aggressive, environmentally independent PCa may be a result of poor stromal ecology, supporting the concept that purely tumour epithelium-centric metrics of aggressiveness may be incomplete and that incorporating markers of stromal ecology would improve prognosis.
Figures
References
-
- Simon-Assmann P, Spenle C, Lefebvre O & Kedinger M The role of the basement membrane as a modulator of intestinal epithelial-mesenchymal interactions. Prog. Mol. Biol. Transl. Sci 96, 175–206 (2010). - PubMed
-
- Parmar H & Cunha GR Epithelial-stromal interactions in the mouse and human mammary gland in vivo. Endocr. Relat. Cancer 11, 437–458 (2004). - PubMed
-
- Sugimoto H, Mundel TM, Kieran MW & Kalluri R Identification of fibroblast heterogeneity in the tumor microenvironment. Cancer Biol. Ther 5, 1640–1646 (2006). - PubMed
-
- Orimo A. et al. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell 121, 335–348 (2005). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
