An official website of the United States government
The .gov means it’s official.
Federal government websites often end in .gov or .mil. Before
sharing sensitive information, make sure you’re on a federal
government site.
The site is secure.
The https:// ensures that you are connecting to the
official website and that any information you provide is encrypted
and transmitted securely.
1 State Key Laboratory of Cancer Biology, Department of Pathology, Xijing Hospital and School of Basic Medicine, Fourth Military Medical University, Xi'an, China.
2 Department of Urology, Xijing Hospital, Fourth Military Medical University, Xi'an, China.
3 Shanghai Key Laboratory of New Drug Design, State Key Laboratory of Bioreactor Engineering, School of Pharmacy, East China University of Science and Technology, Shanghai, China.
4 Cancer Research Institute, Affiliated Hospital of Guilin Medical University, Guilin, China.
5 Department of Biochemistry and Molecular Biology, School of Basic Medicine, Huazhong University of Science and Technology, Wuhan, China.
6 Department of Radiation Oncology and.
7 State Key Laboratory of Cancer Biology, National Clinical Research Center for Digestive Diseases, Xijing Hospital, Fourth Military Medical University, Xi'an, China.
8 Department of Pathology, Second Hospital, Guizhou University of Traditional Chinese Medicine, Guiyang, China.
9 Institute of Pathology and Southwest Cancer Center, Southwest Hospital, Third Military Medical University, Chongqing, China.
10 State Key Laboratory of Cancer Biology, Department of Biochemistry and Molecular Biology, School of Basic Medicine, and.
11 Key Laboratory of Gastrointestinal Pharmacology of Chinese Materia Medica of the State Administration of Traditional Chinese Medicine, School of Pharmacy, Fourth Military Medical University, Xi'an, China.
1 State Key Laboratory of Cancer Biology, Department of Pathology, Xijing Hospital and School of Basic Medicine, Fourth Military Medical University, Xi'an, China.
2 Department of Urology, Xijing Hospital, Fourth Military Medical University, Xi'an, China.
3 Shanghai Key Laboratory of New Drug Design, State Key Laboratory of Bioreactor Engineering, School of Pharmacy, East China University of Science and Technology, Shanghai, China.
4 Cancer Research Institute, Affiliated Hospital of Guilin Medical University, Guilin, China.
5 Department of Biochemistry and Molecular Biology, School of Basic Medicine, Huazhong University of Science and Technology, Wuhan, China.
6 Department of Radiation Oncology and.
7 State Key Laboratory of Cancer Biology, National Clinical Research Center for Digestive Diseases, Xijing Hospital, Fourth Military Medical University, Xi'an, China.
8 Department of Pathology, Second Hospital, Guizhou University of Traditional Chinese Medicine, Guiyang, China.
9 Institute of Pathology and Southwest Cancer Center, Southwest Hospital, Third Military Medical University, Chongqing, China.
10 State Key Laboratory of Cancer Biology, Department of Biochemistry and Molecular Biology, School of Basic Medicine, and.
11 Key Laboratory of Gastrointestinal Pharmacology of Chinese Materia Medica of the State Administration of Traditional Chinese Medicine, School of Pharmacy, Fourth Military Medical University, Xi'an, China.
Esophageal squamous cell carcinoma (ESCC) is one of the most aggressive cancers and is highly resistant to current treatments. ESCC harbors a subpopulation of cells exhibiting cancer stem-like cell (CSC) properties that contribute to therapeutic resistance including radioresistance, but the molecular mechanisms in ESCC CSCs are currently unknown. Here, we report that ribosomal S6 protein kinase 4 (RSK4) plays a pivotal role in promoting CSC properties and radioresistance in ESCC. RSK4 was highly expressed in ESCC CSCs and associated with radioresistance and poor survival in patients with ESCC. RSK4 was found to be a direct downstream transcriptional target of ΔNp63α, the main p63 isoform, which is frequently amplified in ESCC. RSK4 activated the β-catenin signaling pathway through direct phosphorylation of GSK-3β at Ser9. Pharmacologic inhibition of RSK4 effectively reduced CSC properties and improved radiosensitivity in both nude mouse and patient-derived xenograft models. Collectively, our results strongly suggest that the ΔNp63α/RSK4/GSK-3β axis plays a key role in driving CSC properties and radioresistance in ESCC, indicating that RSK4 is a promising therapeutic target for ESCC treatment.
Conflict of interest: The authors have declared that no conflict of interest exists.
Figures
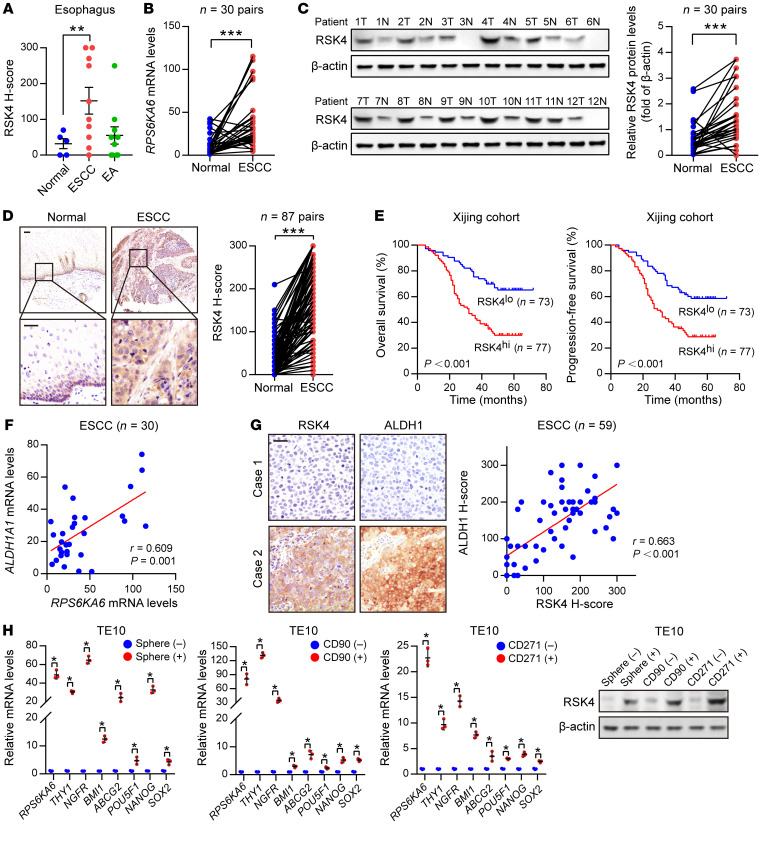
Figure 1. RSK4 is highly expressed in…
Figure 1. RSK4 is highly expressed in ESCC CSCs.
( A ) RSK4 protein was…
Figure 1. RSK4 is highly expressed in ESCC CSCs.
(A) RSK4 protein was highly expressed in ESCC rather than in esophageal adenocarcinoma (EA) compared with expression in corresponding nontumor tissues. Representative IHC images are shown in Supplemental Figure 1A. (B) mRNA levels of RPS6KA6 in 30 pairs of ESCC samples and adjacent nontumor tissues were determined by real-time PCR. GAPDH was used as a loading control. (C) Western blot analysis and quantification of RSK4 expression in ESCC tumor tissues (T) and adjacent nontumor tissues (N) from 30 patients. The results for the other samples are presented in Supplemental Figure 1B. Protein expression was normalized to β-actin levels. (D) Representative IHC images and H-score of RSK4 protein expression in ESCC tumor tissues and adjacent nontumor tissues. Scale bars: 100 μm. (E) Kaplan-Meier estimation of ESCC OS and PFS based on the RSK4 expression levels in the Xijing cohort. (F) Correlation between RPS6KA6 and ALDH1A1 mRNA expression in 30 ESCC patients. (G) Representative IHC images of RSK4 and ALDH1 protein expression in patients with ESCC from the Xijing cohort. Scale bars: 100 μm. Correlation of IHC data on RSK4 and ALDH1 protein expression in 59 ESCC patients. (H) RSK4 was preferentially expressed in tumor spheres compared with nonspheres, and elevated RSK4 expression was detected in CD90+- or CD271+-enriched cell populations compared with the CD90− or CD271− cell subsets as assessed by real-time PCR (n = 3 independent experiments) and immunoblotting. Data represent the mean ± SD. *P < 0.05, **P < 0.01, and ***P < 0.001. Differences were tested using a paired (B–D) and unpaired (H) 2-sided Student’s t test, 1-way ANOVA with Tukey’s post hoc test (A), and log-rank test (E). The correlation was determined by Pearson’s correlation test (F and G).
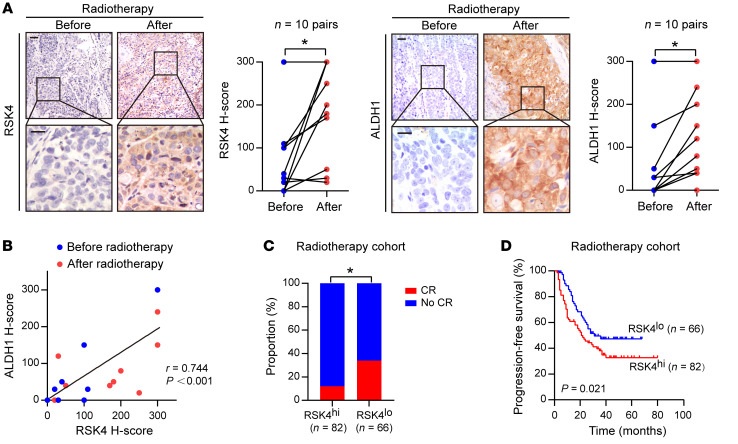
Figure 2. RSK4 is closely linked with…
Figure 2. RSK4 is closely linked with the radioresistance and poor survival of ESCC patients.
Figure 2. RSK4 is closely linked with the radioresistance and poor survival of ESCC patients.
(A) Representative IHC images and IHC H-scores of RSK4 and ALDH1 protein expression in 10 patients with ESCC before and after receiving radiotherapy. Scale bars: 100 μm. (B) RSK4 protein levels were positively correlated with ALDH1 in these cases. (C) Radiotherapy-treated ESCC patients with higher expression of RSK4 had a lower complete response (CR) ratio. (D) Kaplan-Meier estimation of the PFS curves for 148 patients with ESCC treated with radiotherapy according to RSK4 expression levels in the primary tumor. *P < 0.05. Differences were tested using a paired, 2-sided Student’s t test (A), χ2 test (C), and log-rank test (D). The correlation was determined by Pearson’s correlation test (B).
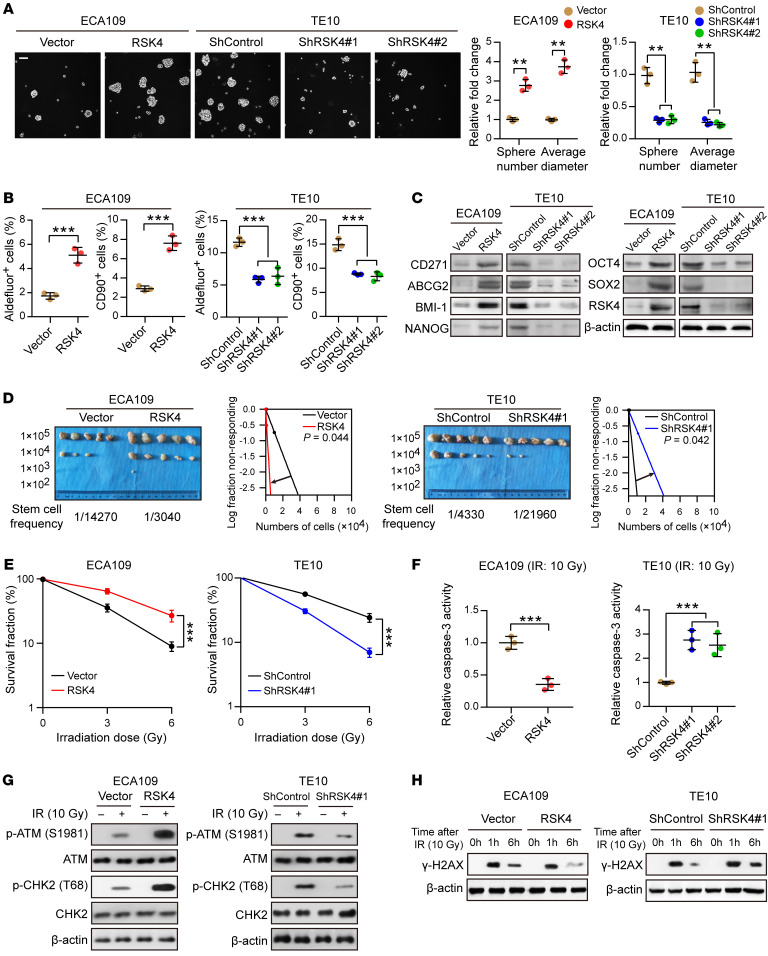
Figure 3. RSK4 promotes the CSC properties…
Figure 3. RSK4 promotes the CSC properties and radioresistance of ESCC cells.
( A )…
Figure 3. RSK4 promotes the CSC properties and radioresistance of ESCC cells.
(A) Tumor sphere formation assay showing that RSK4 overexpression increased the sphere-forming ability of ESCC cells, whereas RSK4 knockdown reduced their sphere-forming ability (n = 3 independent experiments). Scale bar: 100 μm. (B) Flow cytometric analysis showing that RSK4-overexpressing cells had elevated ALDH activity and an increased proportion of CD90+ cells, whereas RSK4-suppressed cells exhibited reduced ALDH activity and a reduced proportion of CD90+ cells (n = 3 independent experiments). (C) Western blot analysis indicating that RSK4 overexpression increased ESCC CSC marker expression, whereas RSK4 knockdown had the opposite effect. (D) Limiting dilution analysis showing higher tumorigenicity of RSK4-overexpressing ECA109 cells in NOD/SCID mice compared with control cells, but RSK4-knockdown TE10 cells had lower tumorigenicity compared with the control group cells (n = 5 mice each). (E) Clonogenic survival assays of ESCC cells with overexpression or knockdown of RSK4 at the indicated irradiation doses (n = 3 independent experiments). (F) Relative caspase-3 activity 24 hours after IR (10 Gy) of ESCC cells with overexpression or knockdown of RSK4 (n = 3 independent experiments). (G) Western blot analysis of phosphorylated and total amounts of the checkpoint proteins ATM and CHK2 from the indicated groups before treatment (–) and 1 hour after 10 Gy IR (+). (H) ESCC cells from the indicated groups were treated with 10 Gy IR and recultured under normal conditions for 1 and 6 hours, and then subjected to Western blot analysis with γ-H2AX antibody. 0h, cells with IR treatment but with no time for DNA repair. Data represent the mean ± SD. **P < 0.01 and ***P < 0.001. Differences were tested using an unpaired, 2-sided Student’s t test (A, B, and D–F).
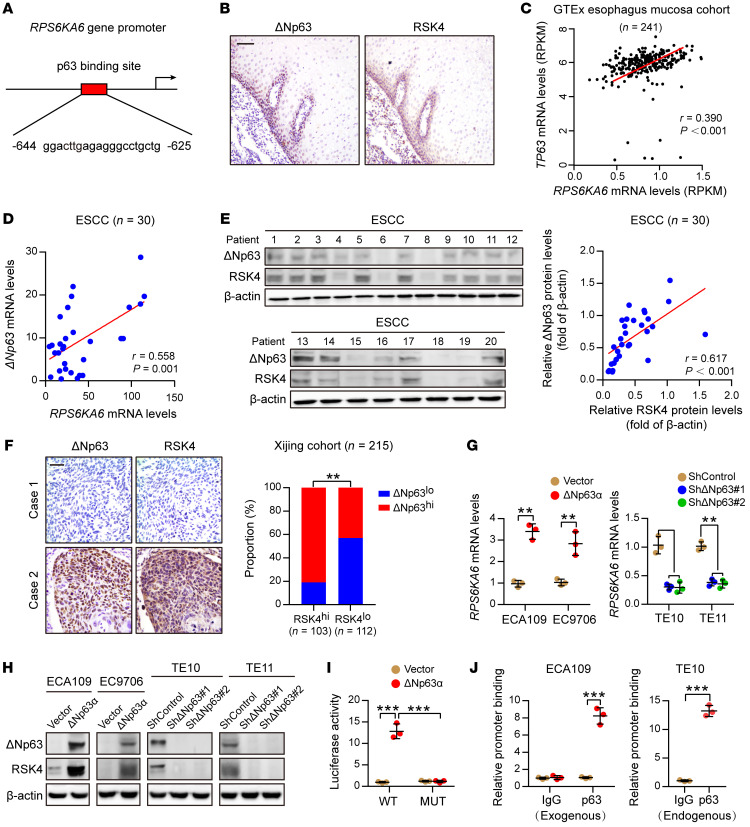
Figure 4. ΔNp63α directly transactivates RSK4 expression in ESCC.
( A ) Predicted binding sites…
Figure 4. ΔNp63α directly transactivates RSK4 expression in ESCC.
(A) Predicted binding sites for p63 in the promoter regions of RPS6KA6. (B) IHC analysis of ΔNp63 and RSK4 proteins showing an identical coexpression pattern in serial sections of the esophageal epithelial basal and suprabasal layers. Scale bar: 50 μm. (C) Correlation between the RPS6KA6 and TP63 mRNA expression patterns in the GTEx esophagus mucosa data set. (D) mRNA levels of ΔNp63 and RPS6KA6 in 30 ESCC samples were determined by real-time PCR. GAPDH was used as a loading control. (E) Expression of ΔNp63 and RSK4 in 30 ESCC samples was detected by Western blotting. The results of other samples are presented in Supplemental Figure 3G. Protein expression was normalized to β-actin levels. (F) Representative IHC images of ΔNp63 and RSK4 protein expression in patients with ESCC. Scale bar: 100 μm. Histograms show the correlation of the IHC data for high or low RSK4 expression relative to the level of ΔNp63. (G) ΔNp63α overexpression upregulated, whereas ΔNp63 silencing reduced, RPS6KA6 mRNA expression in ESCC cells (n = 3 independent experiments). (H) ΔNp63α overexpression upregulated, whereas ΔNp63 silencing reduced, RSK4 protein expression in ESCC cells. (I) ΔNp63α induced reporter activity of the WT RPS6KA6 promoter rather than the p63 responsive element deletion mutant (MUT) promoter, as determined by a luciferase reporter assay in HEK293T cells (n = 3 independent experiments). (J) Exogenous and endogenous ChIP analysis of the interaction between ΔNp63 protein and the RPS6KA6 promoter in ESCC cells (n = 3 independent experiments). Data represent the mean ± SD. **P < 0.01 and ***P < 0.001. Differences were tested using an unpaired, 2-sided Student’s t test (G, I, and J) and a χ2 test (F). The correlation was determined by Pearson’s correlation test (C–E).
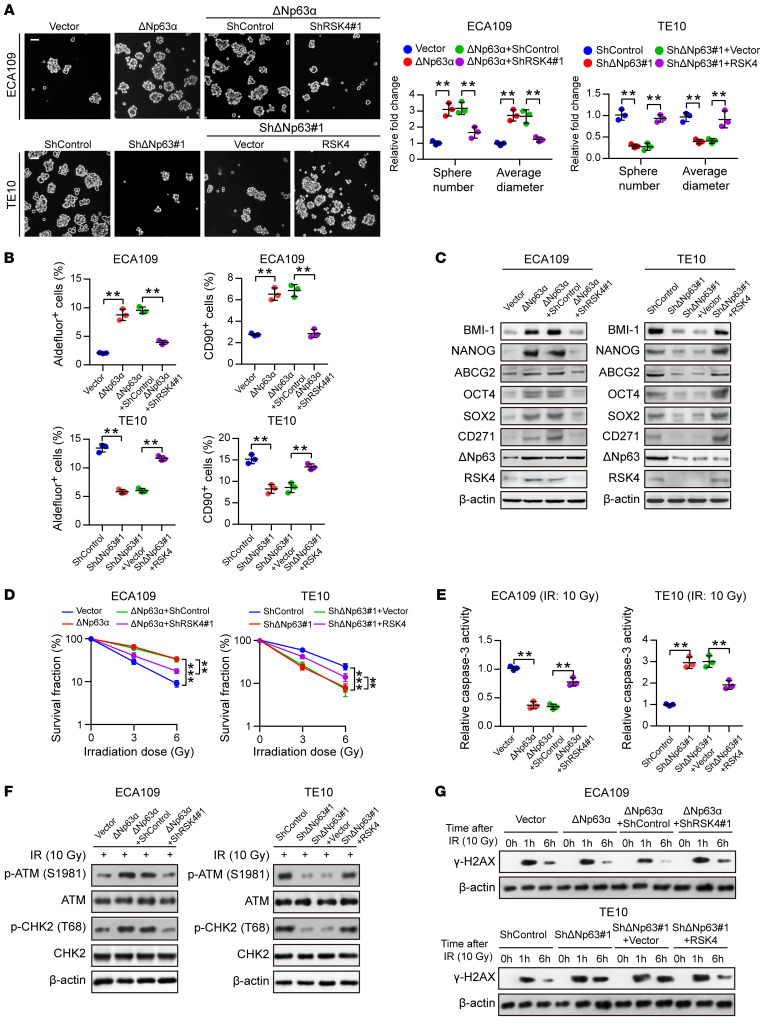
Figure 5. RSK4 is essential for ΔNp63α-mediated…
Figure 5. RSK4 is essential for ΔNp63α-mediated CSC properties and radioresistance of ESCC cells.
( …
Figure 5. RSK4 is essential for ΔNp63α-mediated CSC properties and radioresistance of ESCC cells.
(A) Knockdown of RSK4 abolished ΔNp63α-enhanced sphere formation ability, whereas RSK4 overexpression partially restored the effects of ΔNp63-suppressed sphere formation (n = 3 independent experiments). Scale bars: 100 μm. (B) Flow cytometric analysis of ALDH activity and the proportion of CD90+ cells in ESCC cells from the indicated groups (n = 3 independent experiments). (C) Western blot analysis of ESCC CSC markers in the indicated groups. (D) Clonogenic survival assays of ESCC cells in the indicated groups at IR doses of 0, 3, and 6 Gy (n = 3 independent experiments). (E) Relative caspase-3 activity 24 hours after IR (10 Gy) of ESCC cells in the indicated groups (n = 3 independent experiments). (F) Western blot analysis of phosphorylated and total amounts of the checkpoint proteins ATM and CHK2 from the indicated groups 1 hour after 10 Gy IR. (G) ESCC cells from the indicated groups were treated with 10 Gy IR and recultured under normal conditions for 1 and 6 hours, and then subjected to Western blot analysis with γ-H2AX antibody. 0 h, cells with IR treatment but with no time for DNA repair. Data represent the mean ± SD. **P < 0.01 and ***P < 0.001. Differences were tested using an unpaired, 2-sided Student’s t test (A, B, D, and E).
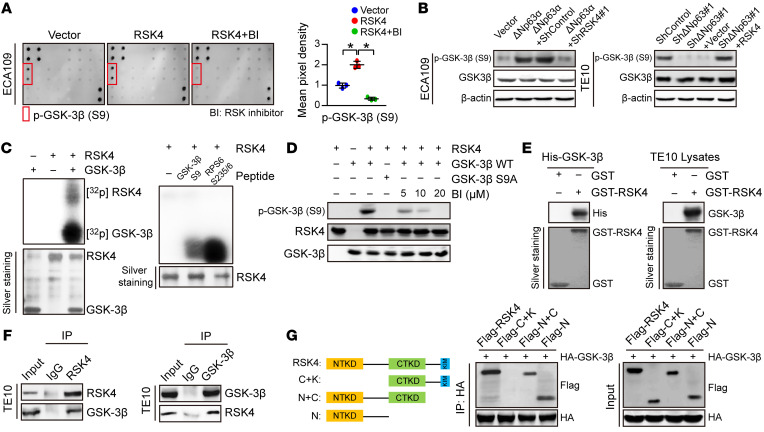
(A) MAPK pathway phosphorylated antibody microarray analysis shows that p–GSK-3β (Ser9) was significantly increased when RSK4 was stably overexpressed and decreased when treated with 10 μM BI-D1870 (an inhibitor of RSK) for 12 hours (n = 3 independent experiments). (B) Western blot analysis showing that downregulation of RSK4 resulted in a reduced level of ΔNp63α-induced p–GSK-3β (Ser9) (S9) (left), whereas RSK4 overexpression partially reversed the reduction in phosphorylation levels through ΔNp63 knockdown (right). (C) Active RSK4 phosphorylated GSK-3β at Ser9 in vitro in the presence of [γ-32P] ATP as visualized by an autoradiograph. The RPS6 (Ser235/236) peptide was used as a positive control. The input was confirmed by silver staining. (D) Validation of p–GSK-3β (Ser9) phosphorylation levels in an in vitro kinase assay by Western blotting. The levels of p–GSK-3β were inhibited when treated with BI-D1870 for 2 hours. WT GSK-3β and mutant GSK-3β S9A were used as substrates for active RSK4. (E) An in vitro GST-pulldown assay was performed to verify the interaction of RSK4 with purified His–GSK-3β protein (left) or GSK-3β from TE10 cell lysates (right). Retrieved proteins were evaluated by immunoblotting. GST-only protein was used as a negative control. GST fusion proteins were confirmed by silver staining. (F) The interaction of RSK4 and GSK-3β was confirmed by an endogenous co-IP assay in TE10 cells. IgG served as a negative control. (G) Mapping analyses of full-length and truncated RSK4 with representative co-IP assays in HEK293T cells showing that the NTKD of RSK4 was responsible for the interaction with GSK-3β. C, CTKD; K, kinase interaction motif (KIM); N, NTKD. Data represent the mean ± SD. *P < 0.05. Differences were tested using 1-way ANOVA with Tukey’s post hoc test (A).
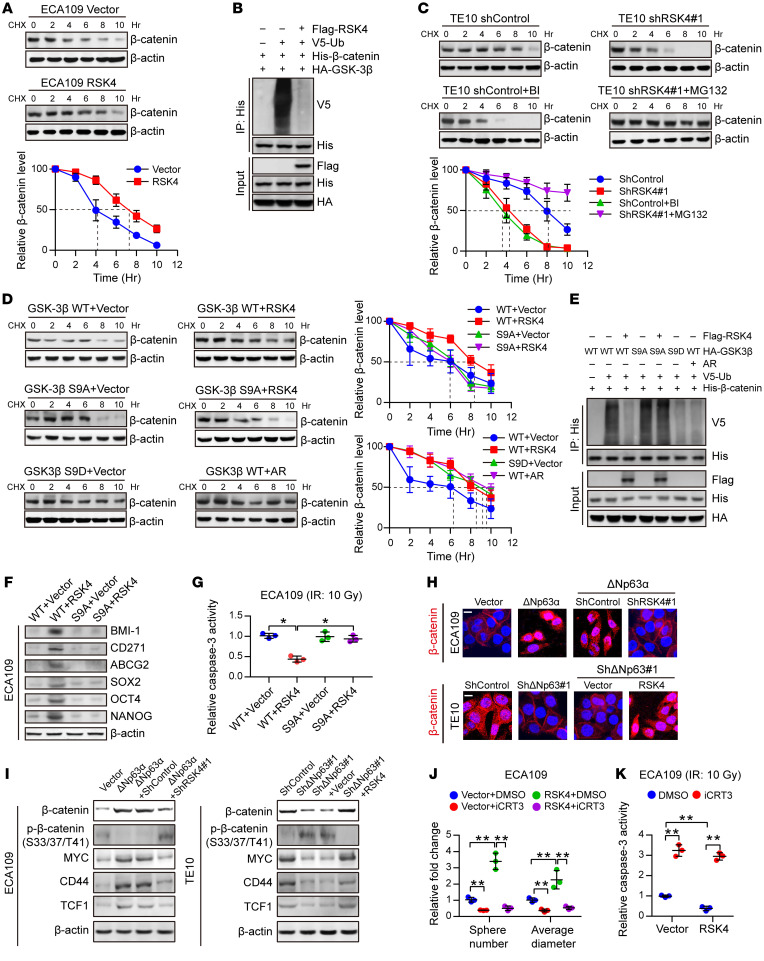
Figure 7. RSK4 activates the Wnt/β-catenin pathway.
Figure 7. RSK4 activates the Wnt/β-catenin pathway.
( A ) Control or RSK4-overexpressing ECA109 cells…
Figure 7. RSK4 activates the Wnt/β-catenin pathway.
(A) Control or RSK4-overexpressing ECA109 cells were incubated with CHX (10 μg/mL) for the indicated durations. High RSK4 expression prolonged the half-life of β-catenin degradation (n = 3 independent experiments). (B) HEK293T cells were transfected with the indicated plasmids, and cell extracts were immunoprecipitated with an anti-His antibody. Ubiquitinated β-catenin was detected by immunoblotting. (C) Control or RSK4-silenced TE10 cells were incubated with CHX (10 μg/mL), CHX plus BI-D1870 (10 μM), or MG132 (10 μM) for the indicated durations. The indicated proteins were detected by immunoblotting (n = 3 independent experiments). (D) HEK293T cells were transfected with the indicated plasmids and incubated with CHX (10 μg/mL) for the indicated durations. The indicated proteins were detected by immunoblotting (n = 3 independent experiments). (E) HEK293T cells were transfected with the indicated plasmids, and cell extracts were immunoprecipitated with an anti-His antibody. Ubiquitinated β-catenin was detected by immunoblotting. (F and G) ECA109 cells were transfected with the indicated plasmids, and then CSC markers (F) and caspase-3 activity after IR (10 Gy) (G) of the indicated groups were detected (n = 3 independent experiments). (H) Representative images of the nuclear localization of β-catenin in ESCC cells from the indicated groups detected using immunofluorescence microscopy. Scale bars: 100 μm. (I) Effects of knockdown or overexpression of RSK4 on the indicated proteins in ΔNp63α-overexpressing or ΔNp63-suppressing ESCC cells. (J and K) Treatment of RSK4-overexpressing cells with 50 μM iCRT3 (an inhibitor of β-catenin signaling) for 24 hours greatly reduced their sphere-forming ability (J) and increased caspase-3 activity after IR (K) (n = 3 independent experiments). BI, BI-D1870; AR, AR-A014418. Data represent the mean ± SD. *P < 0.05 and **P < 0.01. Differences were tested using 1-way ANOVA with Tukey’s post hoc test (G, J, and K).
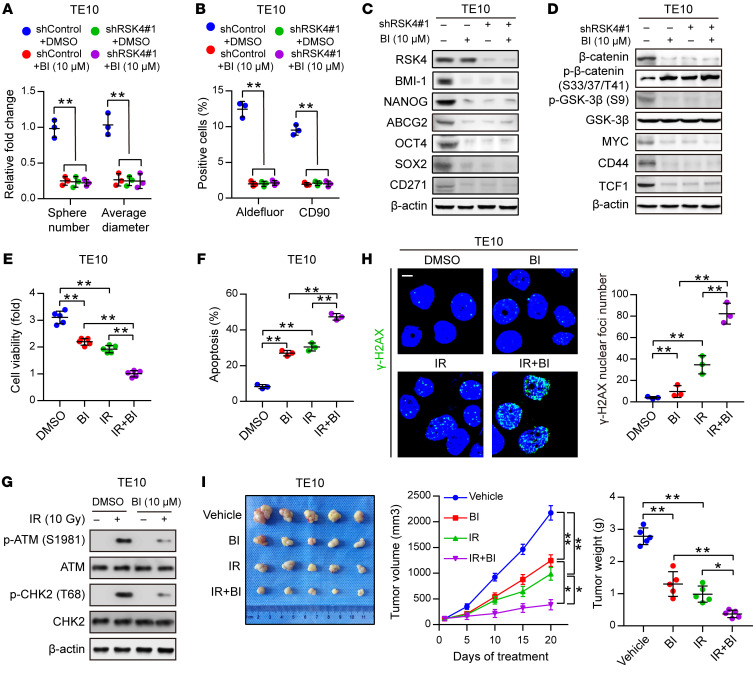
Figure 8. Disruption of the RSK4 pathway…
Figure 8. Disruption of the RSK4 pathway reduces CSC properties and improves the radiosensitivity of…
Figure 8. Disruption of the RSK4 pathway reduces CSC properties and improves the radiosensitivity of ESCC.
(A and B) Sphere formation assay (A) and flow cytometric analysis (B) of TE10 cells treated with BI-D1870 (10 μM), RSK4 knockdown, or both (n = 3 independent experiments). (C) Immunoblot analyses of BMI-1, NANOG, ABCG2, OCT4, SOX2, and CD271 in TE10 cells with the indicated treatments. (D) Immunoblot analyses of β-catenin, p–β-catenin (Ser33/37/Thr41, inactive state), p–GSK-3β (Ser9), GSK-3β, MYC, CD44, and TCF1 in TE10 cells with the indicated treatments. (E) Cell viability assay of TE10 cells treated with BI-D1870 (10 μM) for 12 hours, IR (10 Gy), or both (n = 3 independent experiments). (F) FACS analyses of apoptosis of TE10 cells treated with BI-D1870 (10 μM) for 12 hours, IR (10 Gy), or both (n = 3 independent experiments). (G) Western blot analysis of phosphorylated and total amounts of the checkpoint proteins ATM and CHK2 from TE10 cells treated with or without BI-D1870 (10 μM) before treatment (–) and 1 hour after 10 Gy IR (+). (H) Immunofluorescence staining of γ-H2AX in TE10 cells with the indicated treatments (n = 3 independent experiments). Scale bar: 100 μm. (I) ESCC-derived xenografts in mice treated with vehicle control, BI-D1870 (50 mg/kg/day, i.p. injection), and/or IR (5 Gy, twice). The growth curve of tumor size and average tumor weight are presented (n = 5 mice each). Data represent the mean ± SD. *P < 0.05 and **P < 0.01. Differences were tested using 1-way ANOVA with Tukey’s post hoc test (A, B, E, F, H, and I).
Figure 9. BI-D1870 inhibits tumor growth in…
Figure 9. BI-D1870 inhibits tumor growth in ESCC PDXs.
( A ) Tumor volume growth…
Figure 9. BI-D1870 inhibits tumor growth in ESCC PDXs.
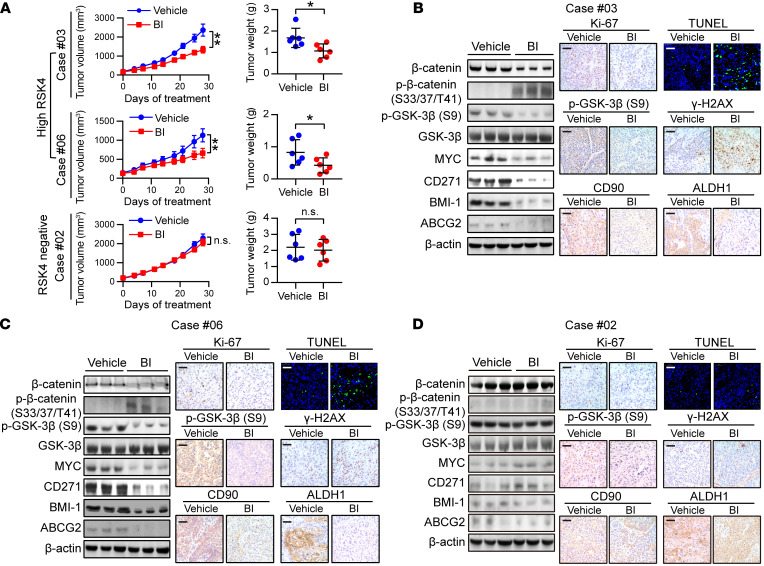
(A) Tumor volume growth curve and tumor weight of PDX mice with different RSK4 expression levels treated with BI-D1870 (50 mg/kg/day, i.p. injection) or vehicle for 28 days (n = 6 mice each). (B–D) Immunoblotting, IHC, and terminal deoxynucleotidyl transferase–mediated dUTP labeling (TUNEL) analyses of the indicated markers in PDX tumor numbers 3 (B), 6 (C) and 2 (D) treated with BI-D1870 (50 mg/kg/day, i.p. injection) or vehicle. Scale bars: 100 μm. Data represent the mean ± SD. *P < 0.05 and **P < 0.01. Differences were tested using an unpaired, 2-sided Student’s t test (A).
Figure 10. BI-D1870 improves the therapeutic efficacy…
Figure 10. BI-D1870 improves the therapeutic efficacy of radiotherapy in ESCC PDXs.
( A )…
Figure 10. BI-D1870 improves the therapeutic efficacy of radiotherapy in ESCC PDXs.
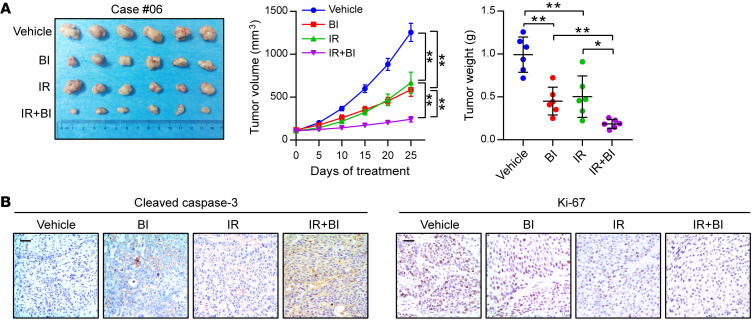
(A) PDX tumor number 6 treated with the vehicle control, BI-D1870 (50 mg/kg/day, i.p. injection), and/or IR (5 Gy, twice). The growth curve of the tumor size and average tumor weight are presented (n = 6 mice each). (B) IHC analyses of cleaved caspase-3 and Ki-67 in the indicated groups. Scale bars: 100 μm. Data represent the mean ± SD. *P < 0.05 and **P < 0.01. Differences were tested using 1-way ANOVA with Tukey’s post hoc test (A).
Abnet CC, Arnold M, Wei WQ. Epidemiology of esophageal squamous cell carcinoma. Gastroenterology. 2018;154(2):360–373. doi: 10.1053/j.gastro.2017.08.023.
-
DOI
-
PMC
-
PubMed
Chen W, et al. Cancer statistics in China, 2015. CA Cancer J Clin. 2016;66(2):115–132. doi: 10.3322/caac.21338.
-
DOI
-
PubMed
Liang H, Fan JH, Qiao YL. Epidemiology, etiology, and prevention of esophageal squamous cell carcinoma in China. Cancer Biol Med. 2017;14(1):33–41. doi: 10.20892/j.issn.2095-3941.2016.0093.
-
DOI
-
PMC
-
PubMed
van Hagen P, et al. Preoperative chemoradiotherapy for esophageal or junctional cancer. N Engl J Med. 2012;366(22):2074–2084. doi: 10.1056/NEJMoa1112088.
-
DOI
-
PubMed
Plaks V, Kong N, Werb Z. The cancer stem cell niche: how essential is the niche in regulating stemness of tumor cells? Cell Stem Cell. 2015;16(3):225–238. doi: 10.1016/j.stem.2015.02.015.
-
DOI
-
PMC
-
PubMed