

Extracellular Signal-Regulated Kinases Mediate an Autoregulation of GABAB-Receptor-Activated Whole-Cell Current in Locus Coeruleus Neurons
- PMID: 32398643
- PMCID: PMC7217949
- DOI: 10.1038/s41598-020-64292-x
Extracellular Signal-Regulated Kinases Mediate an Autoregulation of GABAB-Receptor-Activated Whole-Cell Current in Locus Coeruleus Neurons
Abstract
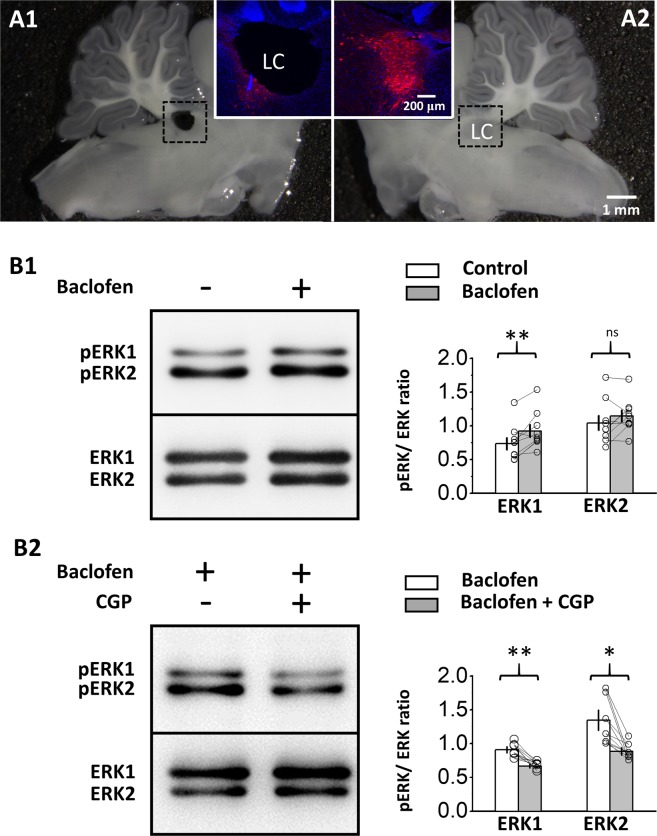
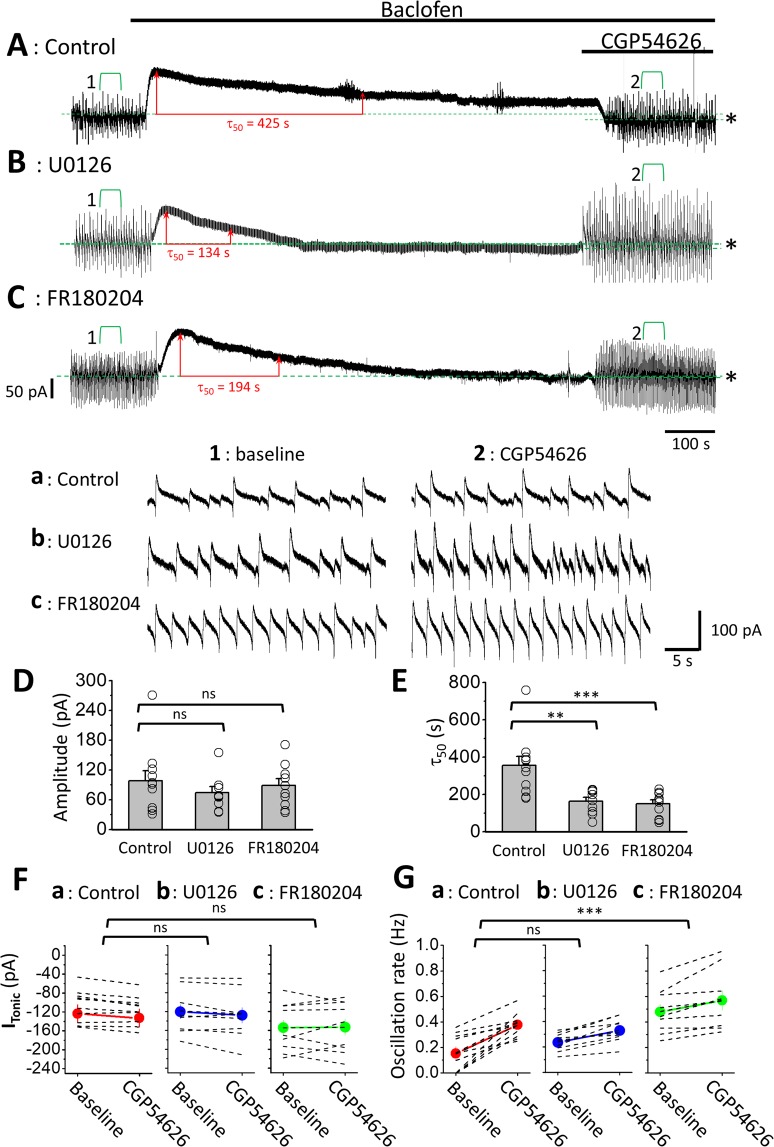
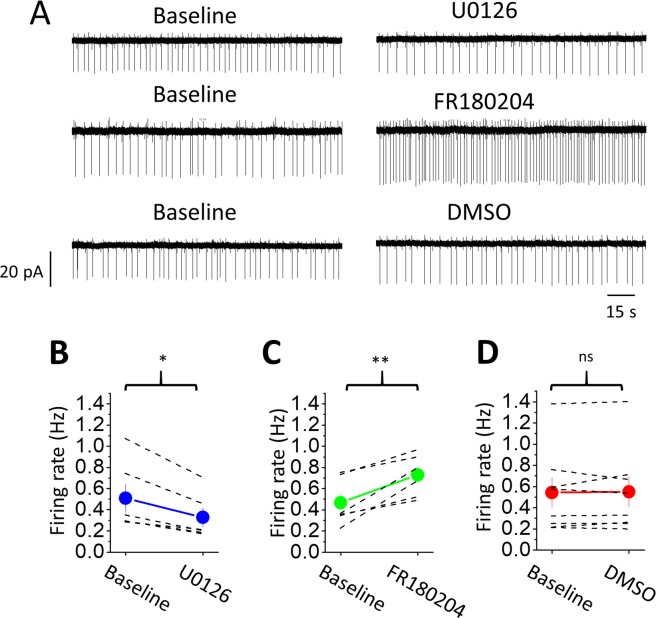
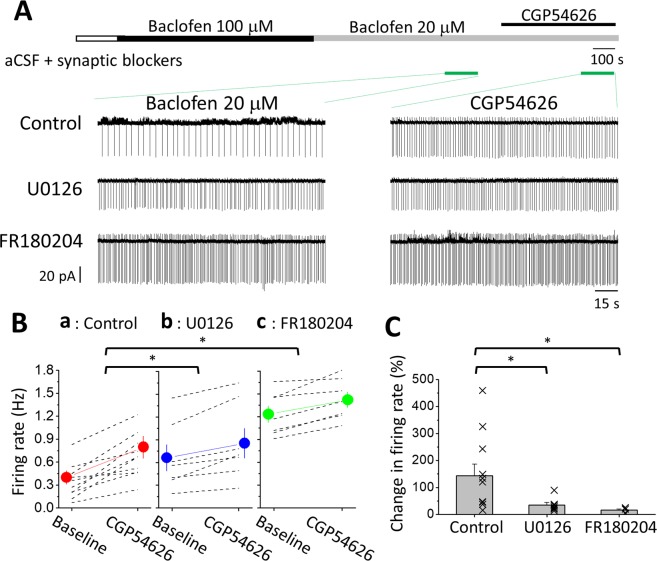
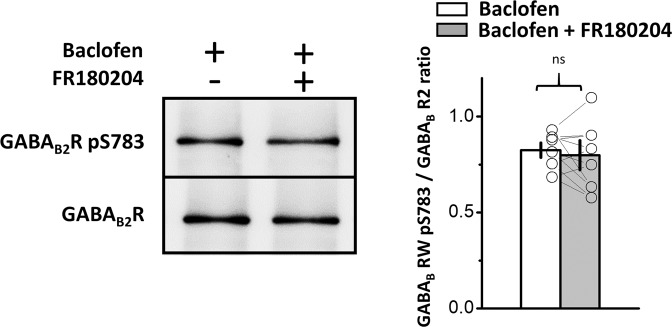
The norepinephrine-releasing neurons in the locus coeruleus (LC) are well known to regulate wakefulness/arousal. They display active firing during wakefulness and a decreased discharge rate during sleep. We have previously reported that LC neurons express large numbers of GABAB receptors (GABABRs) located at peri-/extrasynaptic sites and are subject to tonic inhibition due to the continuous activation of GABABRs by ambient GABA, which is significantly higher during sleep than during wakefulness. In this study, we further showed using western blot analysis that the activation of GABABRs with baclofen could increase the level of phosphorylated extracellular signal-regulated kinase 1 (ERK1) in LC tissue. Recordings from LC neurons in brain slices showed that the inhibition of ERK1/2 with U0126 and FR180204 accelerated the decay of whole-cell membrane current induced by prolonged baclofen application. In addition, the inhibition of ERK1/2 also increased spontaneous firing and reduced tonic inhibition of LC neurons after prolonged exposure to baclofen. These results suggest a new role of GABABRs in mediating ERK1-dependent autoregulation of the stability of GABABR-activated whole-cell current, in addition to its well-known effect on gated potassium channels, to cause a tonic current in LC neurons.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Walker, M.C.&Kullmann, D.M. Tonic GABAA receptor-mediated signaling in epilepsy. In:Jasper’s Basic Mechanisms of the Epilepsies [Internet]. (eds Noebels, J. L., Avoli, M., Rogawski, M. A., Olsen, R. W. & Delgado-Escueta, A. V.) 4th edition. (Bethesda (MD): National Center for Biotechnology Information(US) (2012). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
