

Identification of RAG-like transposons in protostomes suggests their ancient bilaterian origin
- PMID: 32399063
- PMCID: PMC7204232
- DOI: 10.1186/s13100-020-00214-y
Identification of RAG-like transposons in protostomes suggests their ancient bilaterian origin
Abstract
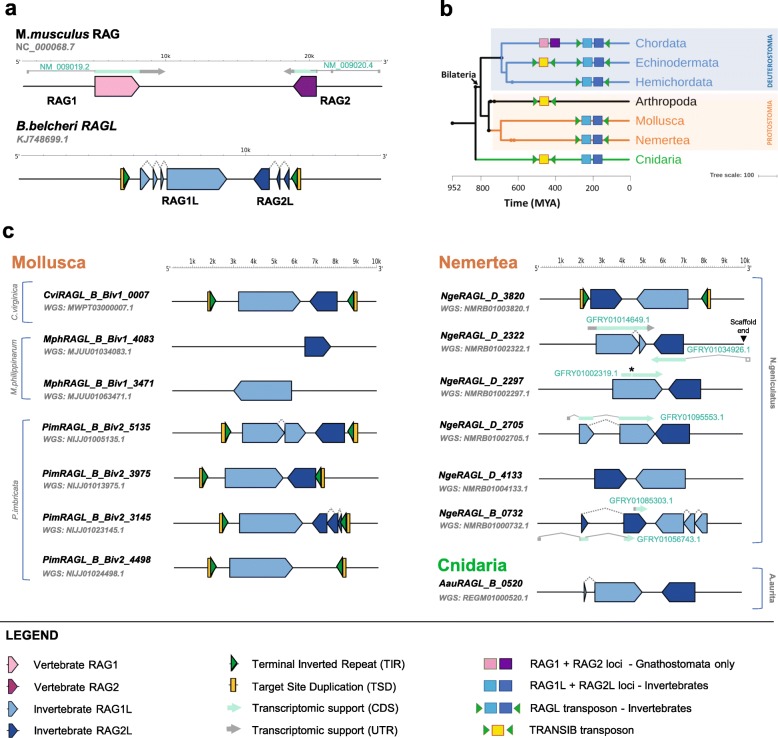
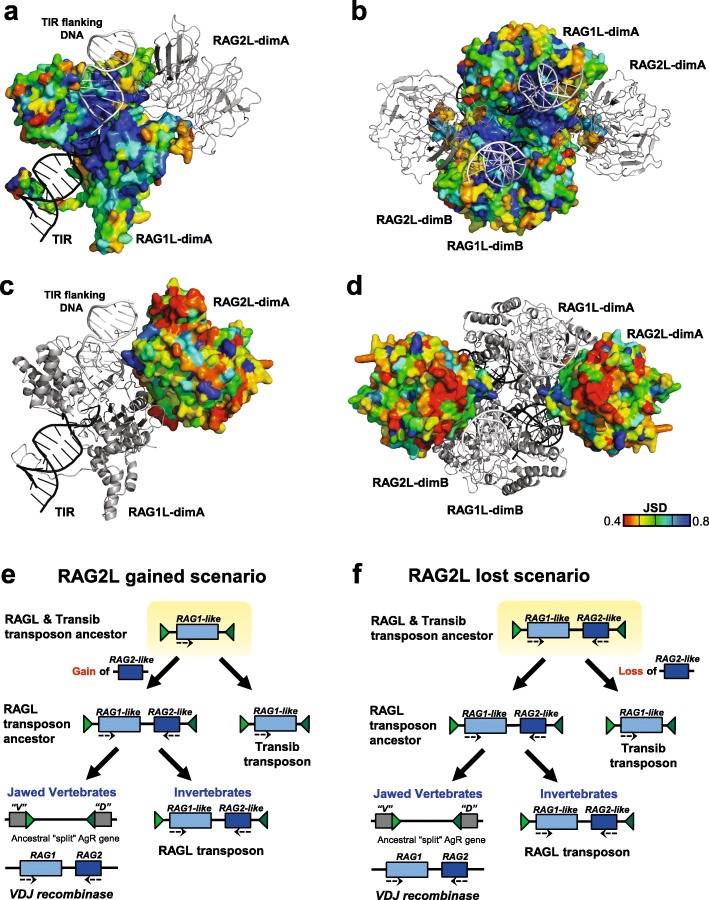
Background: V(D) J recombination is essential for adaptive immunity in jawed vertebrates and is initiated by the RAG1-RAG2 endonuclease. The RAG1 and RAG2 genes are thought to have evolved from a RAGL (RAG-like) transposon containing convergently-oriented RAG1-like (RAG1L) and RAG2-like (RAG2L) genes. Elements resembling this presumptive evolutionary precursor have thus far only been detected convincingly in deuterostomes, leading to the model that the RAGL transposon first appeared in an early deuterostome.
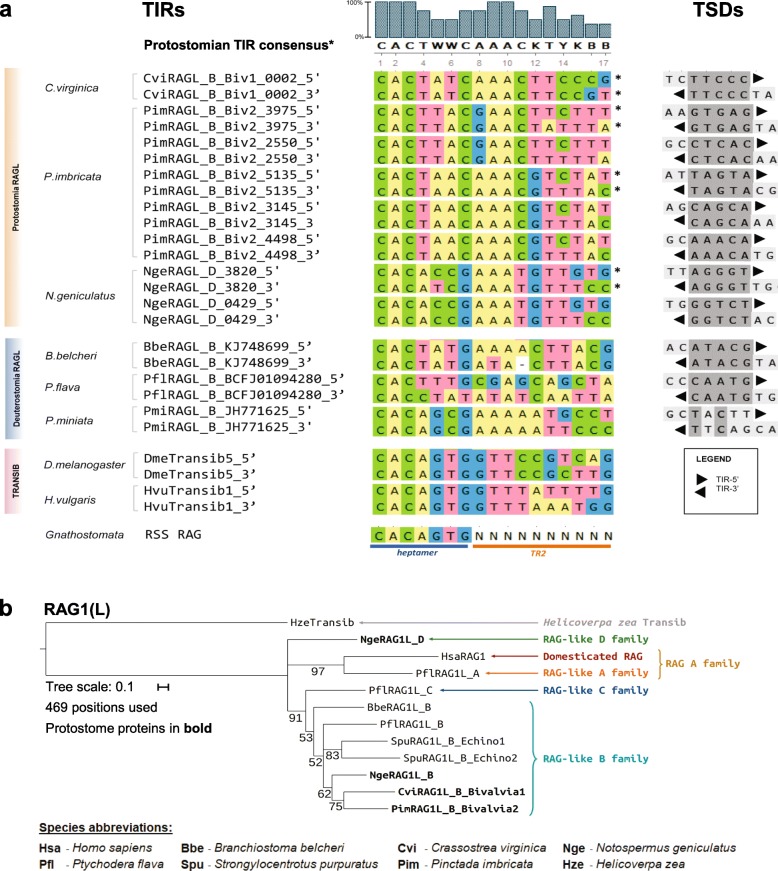
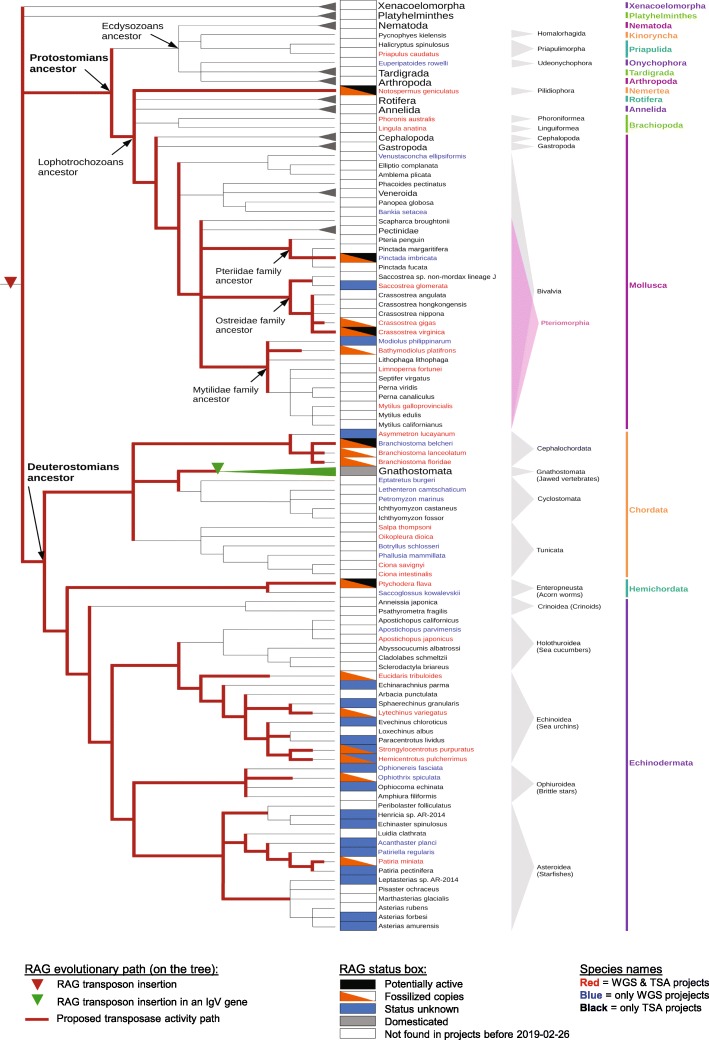
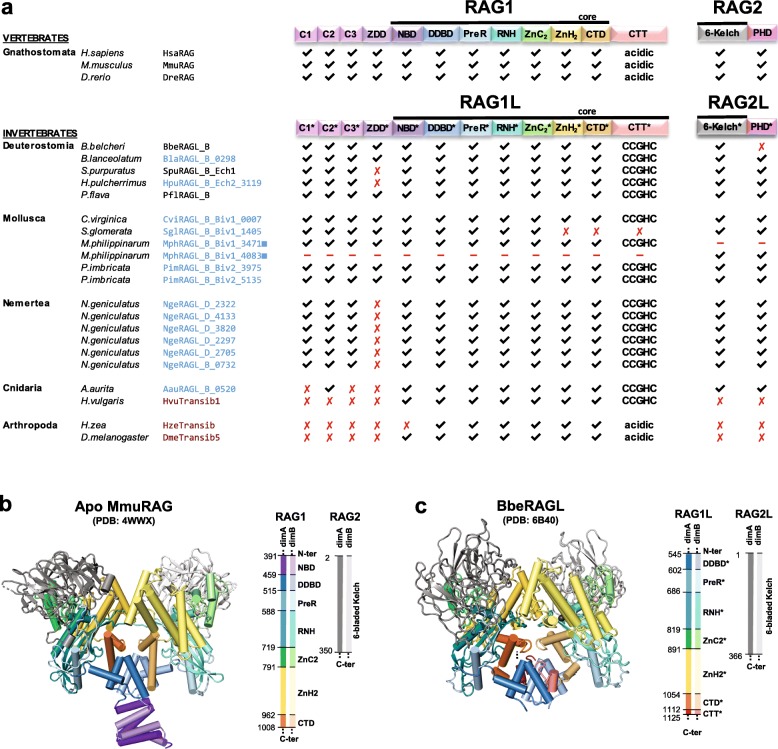
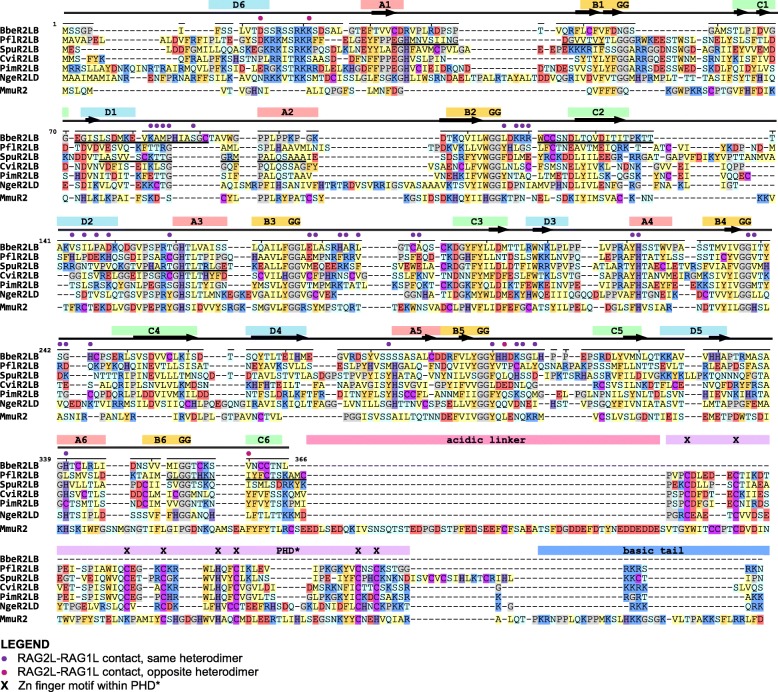
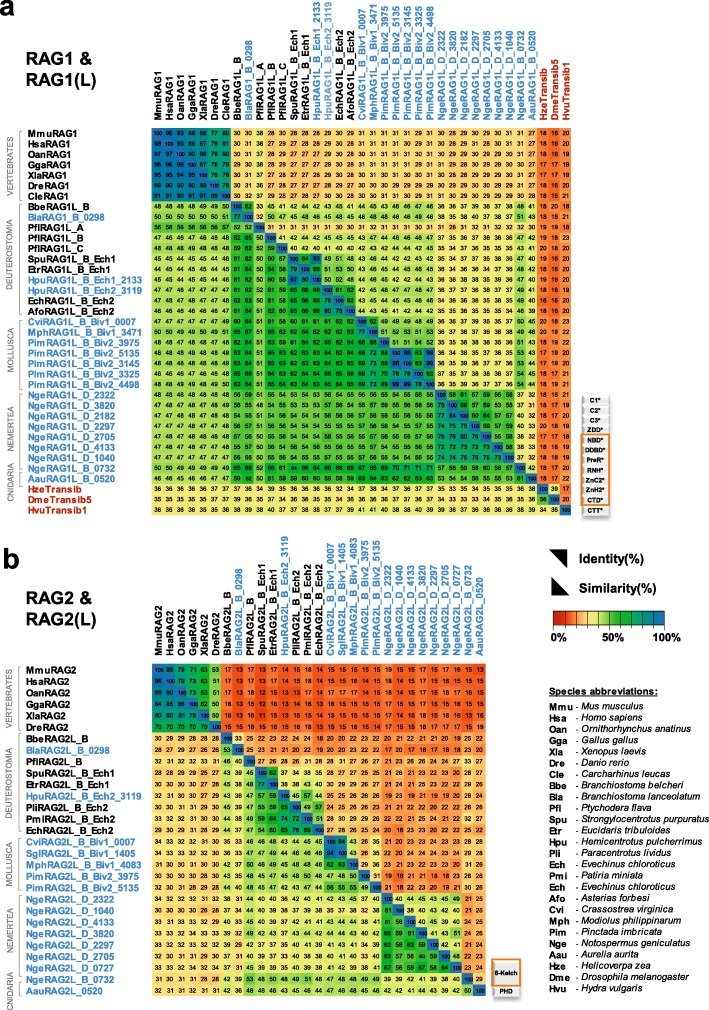
Results: We have identified numerous RAGL transposons in the genomes of protostomes, including oysters and mussels (phylum Mollusca) and a ribbon worm (phylum Nemertea), and in the genomes of several cnidarians. Phylogenetic analyses are consistent with vertical evolution of RAGL transposons within the Bilateria clade and with its presence in the bilaterian ancestor. Many of the RAGL transposons identified in protostomes are intact elements containing convergently oriented RAG1L and RAG2L genes flanked by terminal inverted repeats (TIRs) and target site duplications with striking similarities with the corresponding elements in deuterostomes. In addition, protostome genomes contain numerous intact RAG1L-RAG2L adjacent gene pairs that lack detectable flanking TIRs. Domains and critical active site and structural amino acids needed for endonuclease and transposase activity are present and conserved in many of the predicted RAG1L and RAG2L proteins encoded in protostome genomes.
Conclusions: Active RAGL transposons were present in multiple protostome lineages and many were likely transmitted vertically during protostome evolution. It appears that RAGL transposons were broadly active during bilaterian evolution, undergoing multiple duplication and loss/fossilization events, with the RAGL genes that persist in present day protostomes perhaps constituting both active RAGL transposons and domesticated RAGL genes. Our findings raise the possibility that the RAGL transposon arose earlier in evolution than previously thought, either in an early bilaterian or prior to the divergence of bilaterians and non-bilaterians, and alter our understanding of the evolutionary history of this important group of transposons.
Keywords: Adaptive immunity; Evolution; RAG; Recombination activating genes; Transposon; Transposon molecular domestication.
© The Author(s) 2020.
Conflict of interest statement
Competing interestsThe authors declare that they have no competing interest.
Figures

References
Grants and funding
LinkOut - more resources
Full Text Sources
