

Highly Efficient Gene Editing of Cystic Fibrosis Patient-Derived Airway Basal Cells Results in Functional CFTR Correction
- PMID: 32402246
- PMCID: PMC7335734
- DOI: 10.1016/j.ymthe.2020.04.021
Highly Efficient Gene Editing of Cystic Fibrosis Patient-Derived Airway Basal Cells Results in Functional CFTR Correction
Abstract
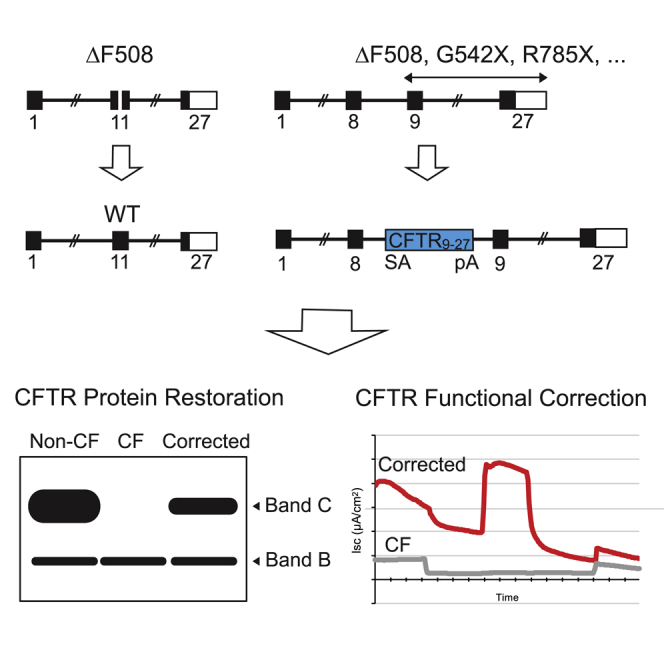
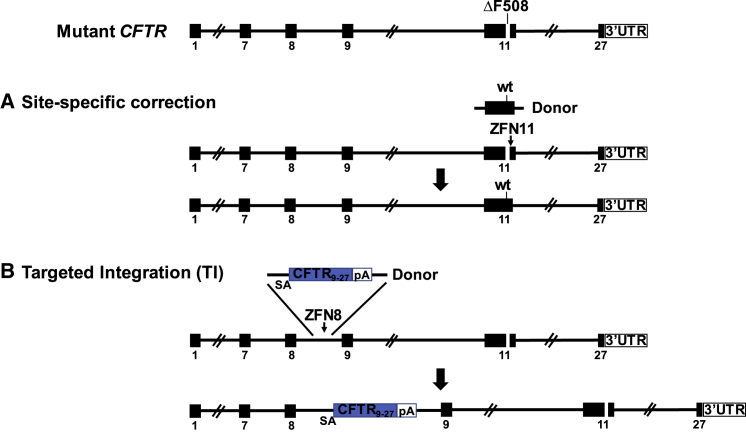
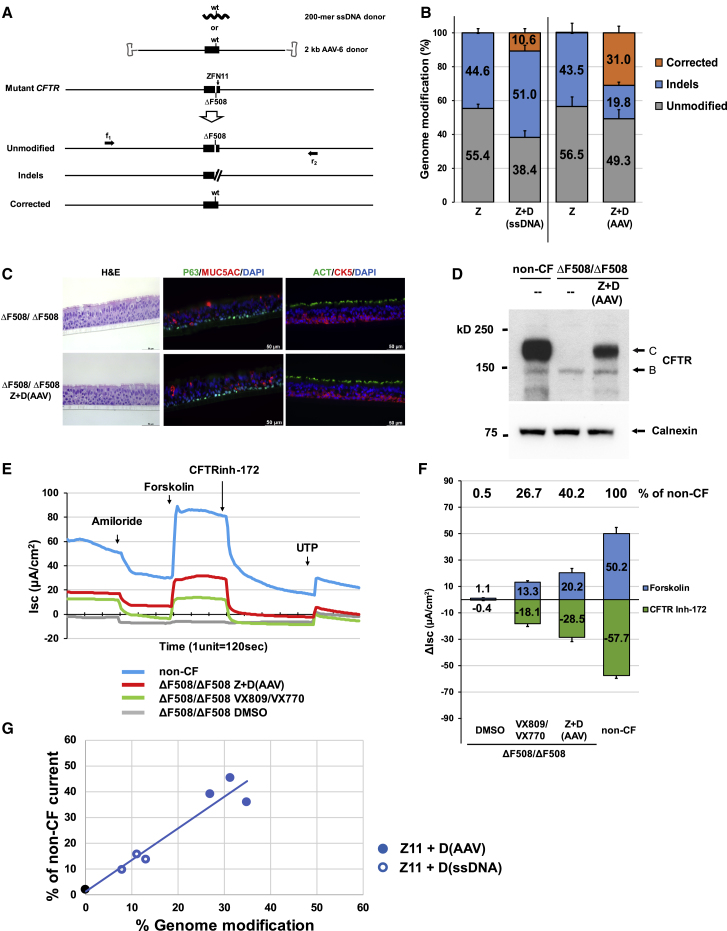
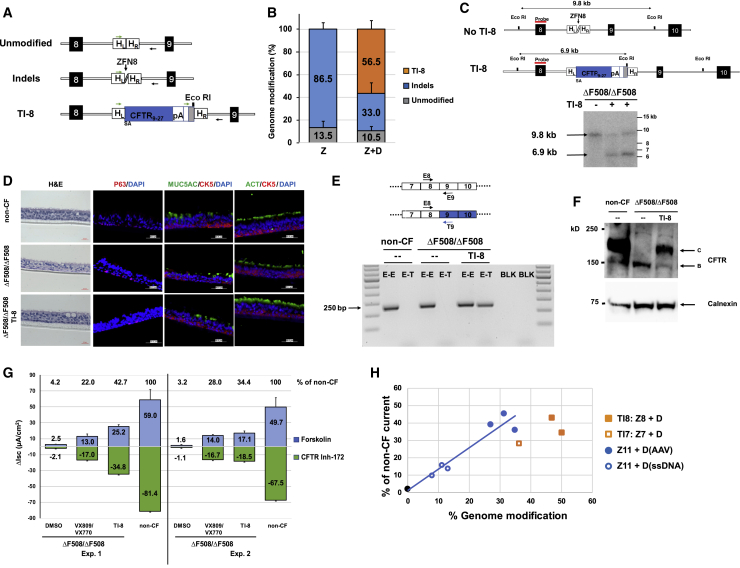
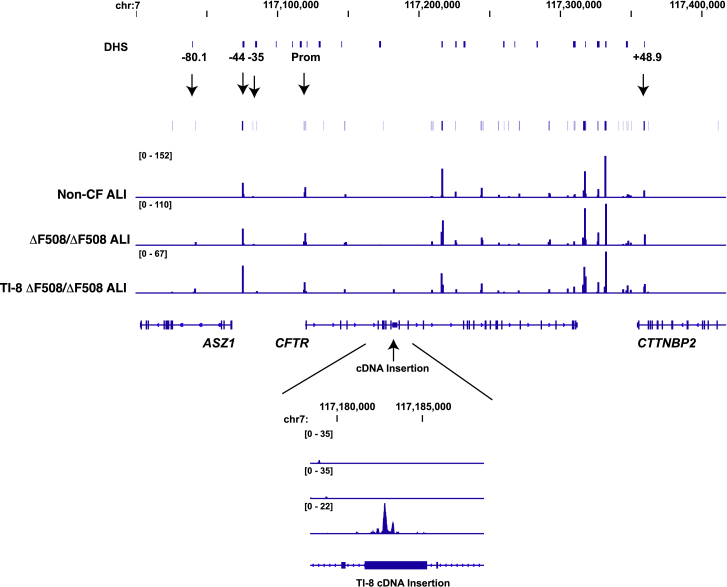
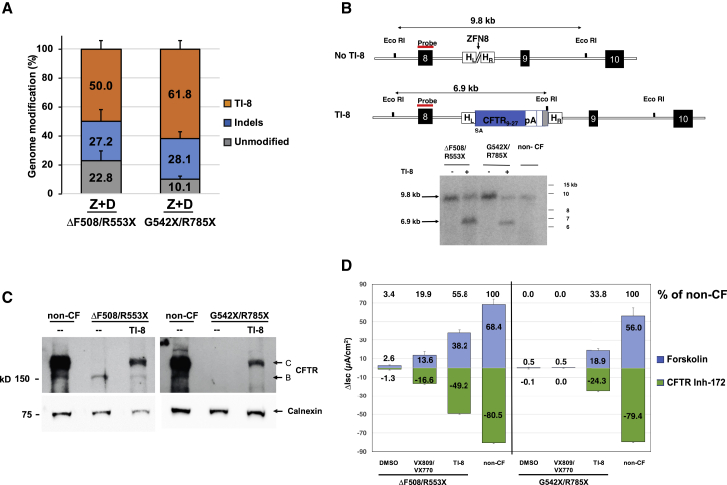
There is a strong rationale to consider future cell therapeutic approaches for cystic fibrosis (CF) in which autologous proximal airway basal stem cells, corrected for CFTR mutations, are transplanted into the patient's lungs. We assessed the possibility of editing the CFTR locus in these cells using zinc-finger nucleases and have pursued two approaches. The first, mutation-specific correction, is a footprint-free method replacing the CFTR mutation with corrected sequences. We have applied this approach for correction of ΔF508, demonstrating restoration of mature CFTR protein and function in air-liquid interface cultures established from bulk edited basal cells. The second is targeting integration of a partial CFTR cDNA within an intron of the endogenous CFTR gene, providing correction for all CFTR mutations downstream of the integration and exploiting the native CFTR promoter and chromatin architecture for physiologically relevant expression. Without selection, we observed highly efficient, site-specific targeted integration in basal cells carrying various CFTR mutations and demonstrated restored CFTR function at therapeutically relevant levels. Significantly, Omni-ATAC-seq analysis revealed minimal impact on the positions of open chromatin within the native CFTR locus. These results demonstrate efficient functional correction of CFTR and provide a platform for further ex vivo and in vivo editing.
Keywords: basal stem cell; cystic fibrosis; gene editing.
Copyright © 2020 The American Society of Gene and Cell Therapy. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Harrison P.T., Hoppe N., Martin U. Gene editing & stem cells. J. Cyst. Fibros. 2018;17:10–16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
