

Activation of Phospholipase C β by Gβγ and Gαq Involves C-Terminal Rearrangement to Release Autoinhibition
- PMID: 32402248
- PMCID: PMC7891876
- DOI: 10.1016/j.str.2020.04.012
Activation of Phospholipase C β by Gβγ and Gαq Involves C-Terminal Rearrangement to Release Autoinhibition
Abstract
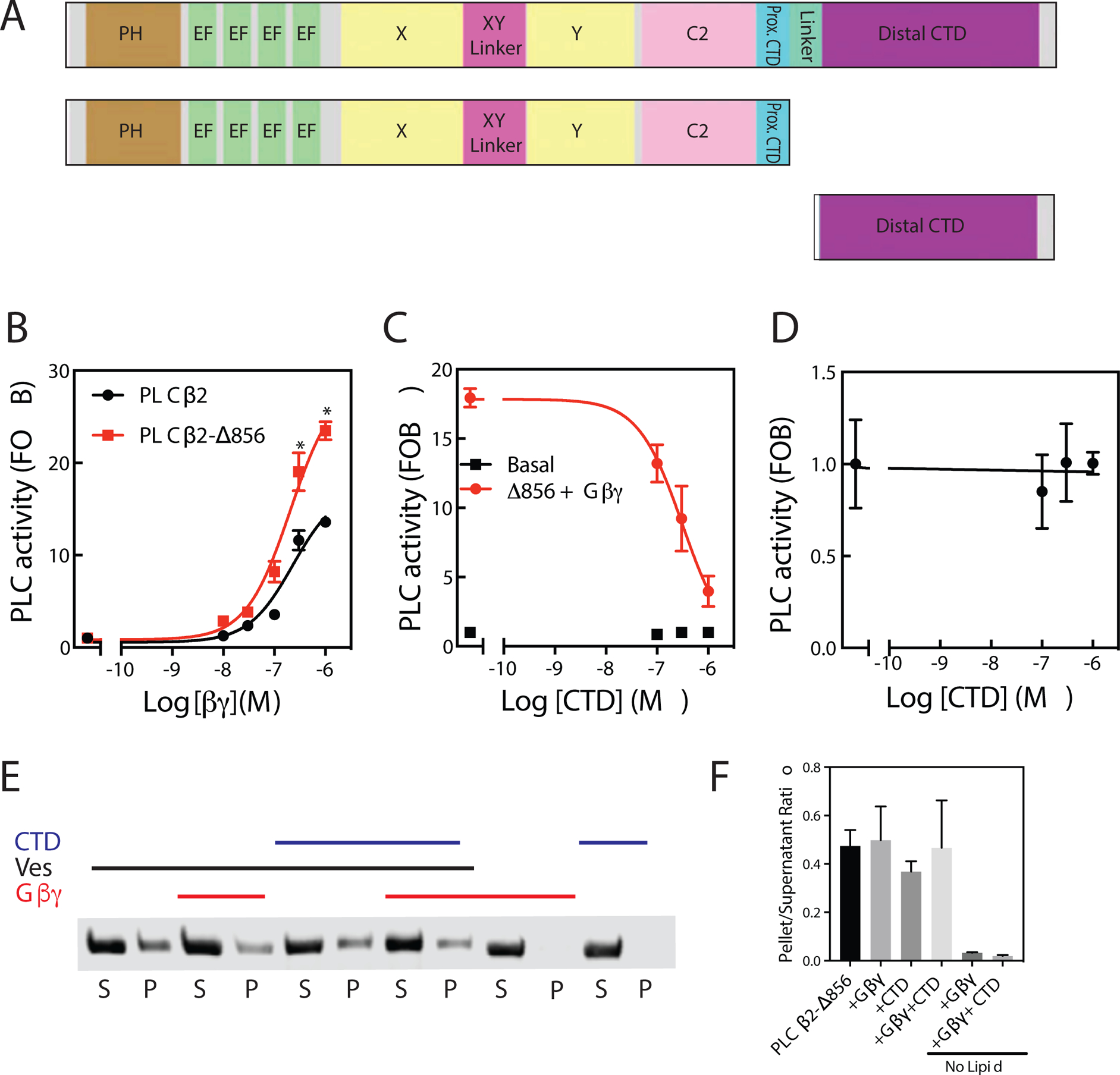
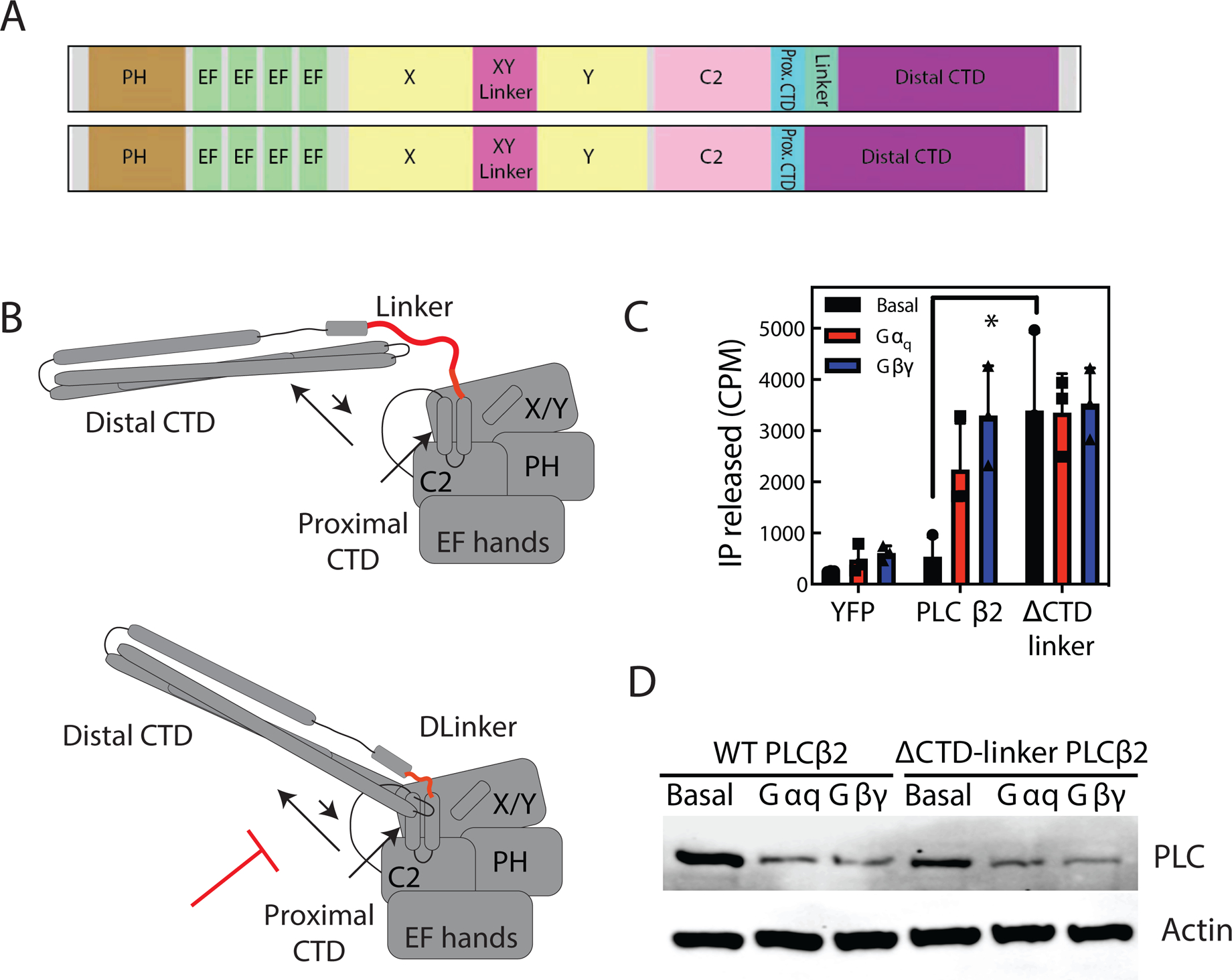
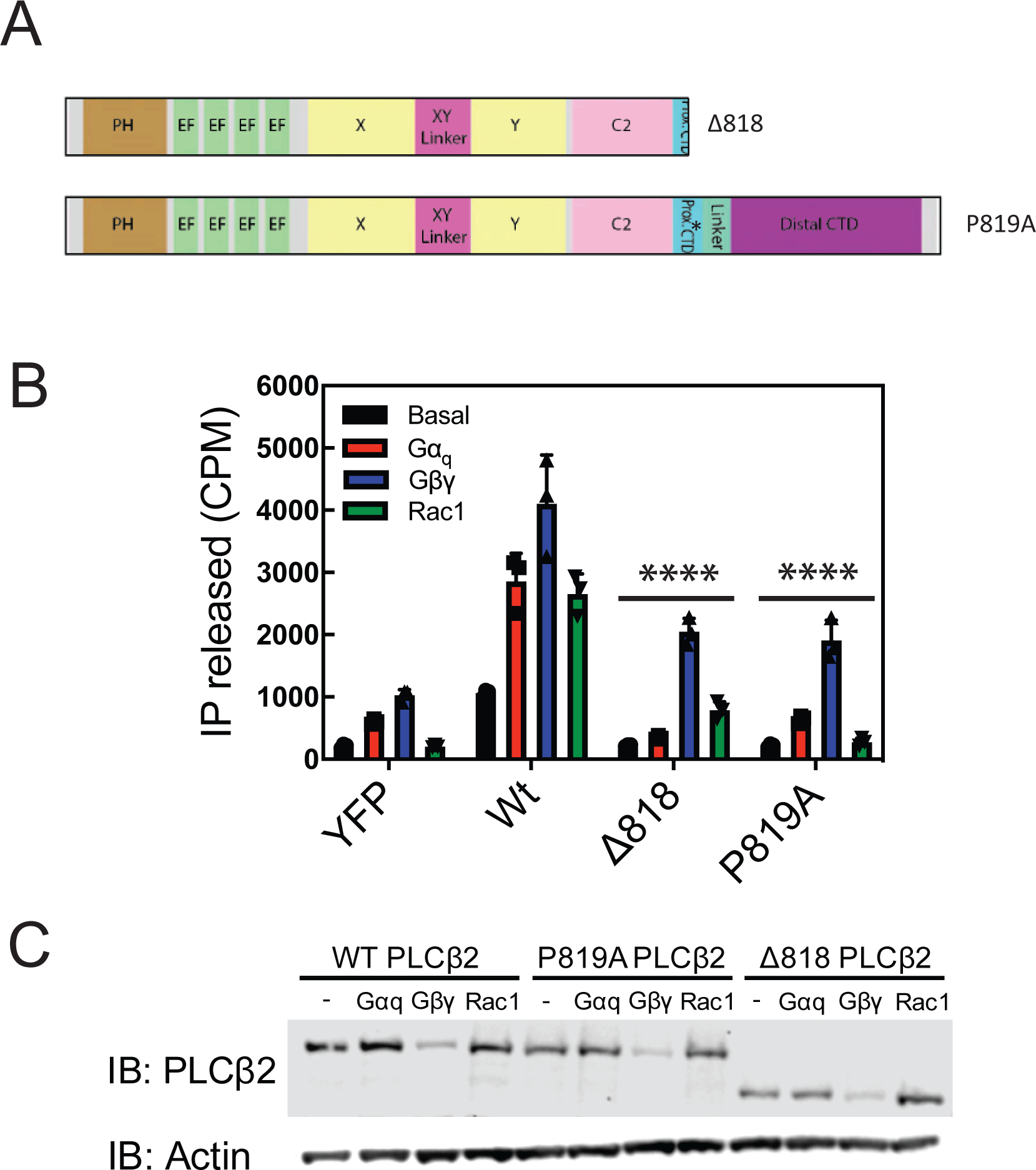
Phospholipase C (PLC) enzymes hydrolyze phosphoinositide lipids to inositol phosphates and diacylglycerol. Direct activation of PLCβ by Gαq and/or Gβγ subunits mediates signaling by Gq and some Gi coupled G-protein-coupled receptors (GPCRs), respectively. PLCβ isoforms contain a unique C-terminal extension, consisting of proximal and distal C-terminal domains (CTDs) separated by a flexible linker. The structure of PLCβ3 bound to Gαq is known, however, for both Gαq and Gβγ; the mechanism for PLCβ activation on membranes is unknown. We examined PLCβ2 dynamics on membranes using hydrogen-deuterium exchange mass spectrometry (HDX-MS). Gβγ caused a robust increase in dynamics of the distal C-terminal domain (CTD). Gαq showed decreased deuterium incorporation at the Gαq binding site on PLCβ. In vitro Gβγ-dependent activation of PLC is inhibited by the distal CTD. The results suggest that disruption of autoinhibitory interactions with the CTD leads to increased PLCβ hydrolase activity.
Keywords: G protein; G-protein-coupled receptor; GPCR; Gαq; Gβγ; HDX-MS; hydrogen-deuterium exchange; lipids; membrane interactions; phospholipase C; protein dynamics; signal transduction.
Copyright © 2020 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
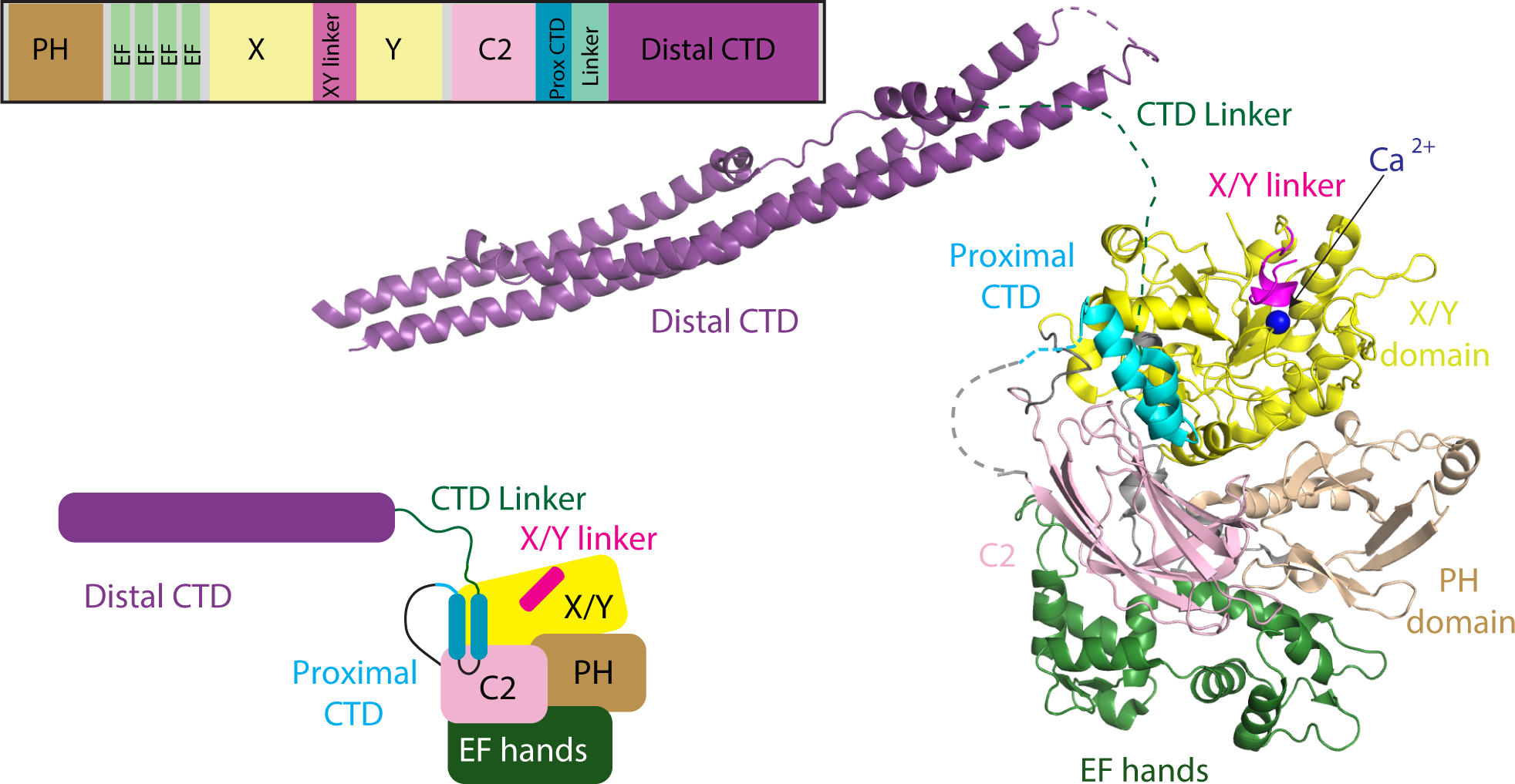
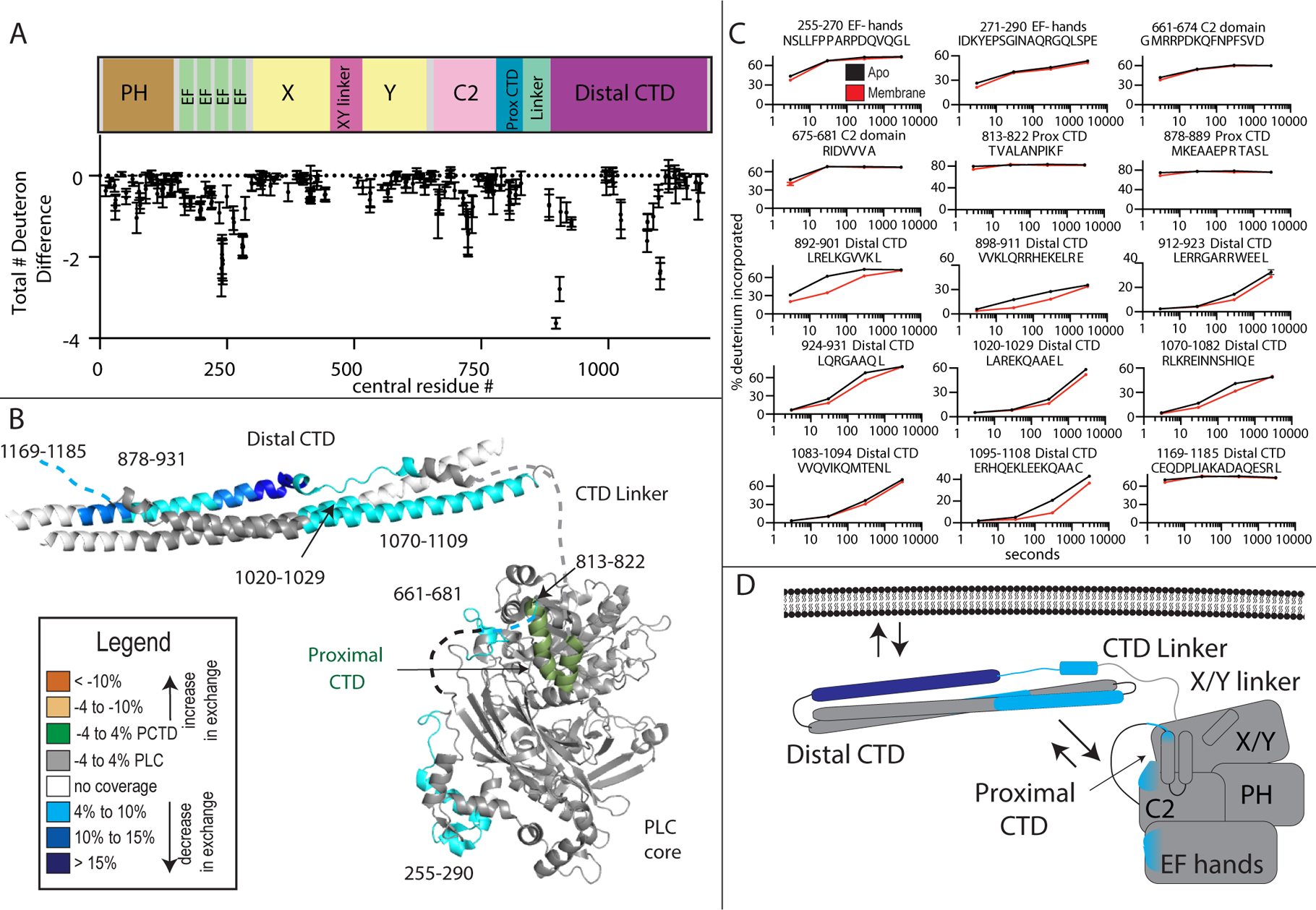
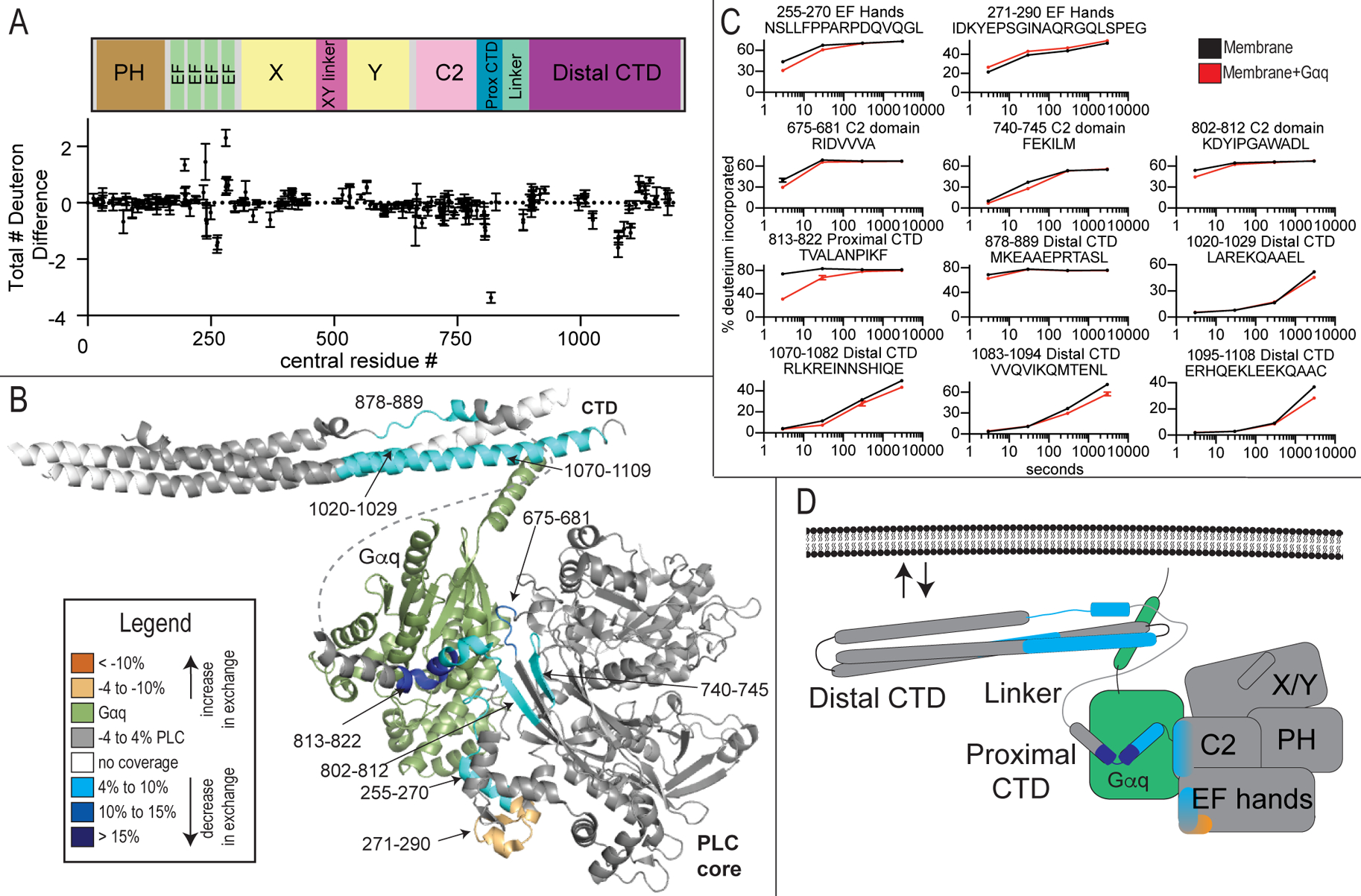
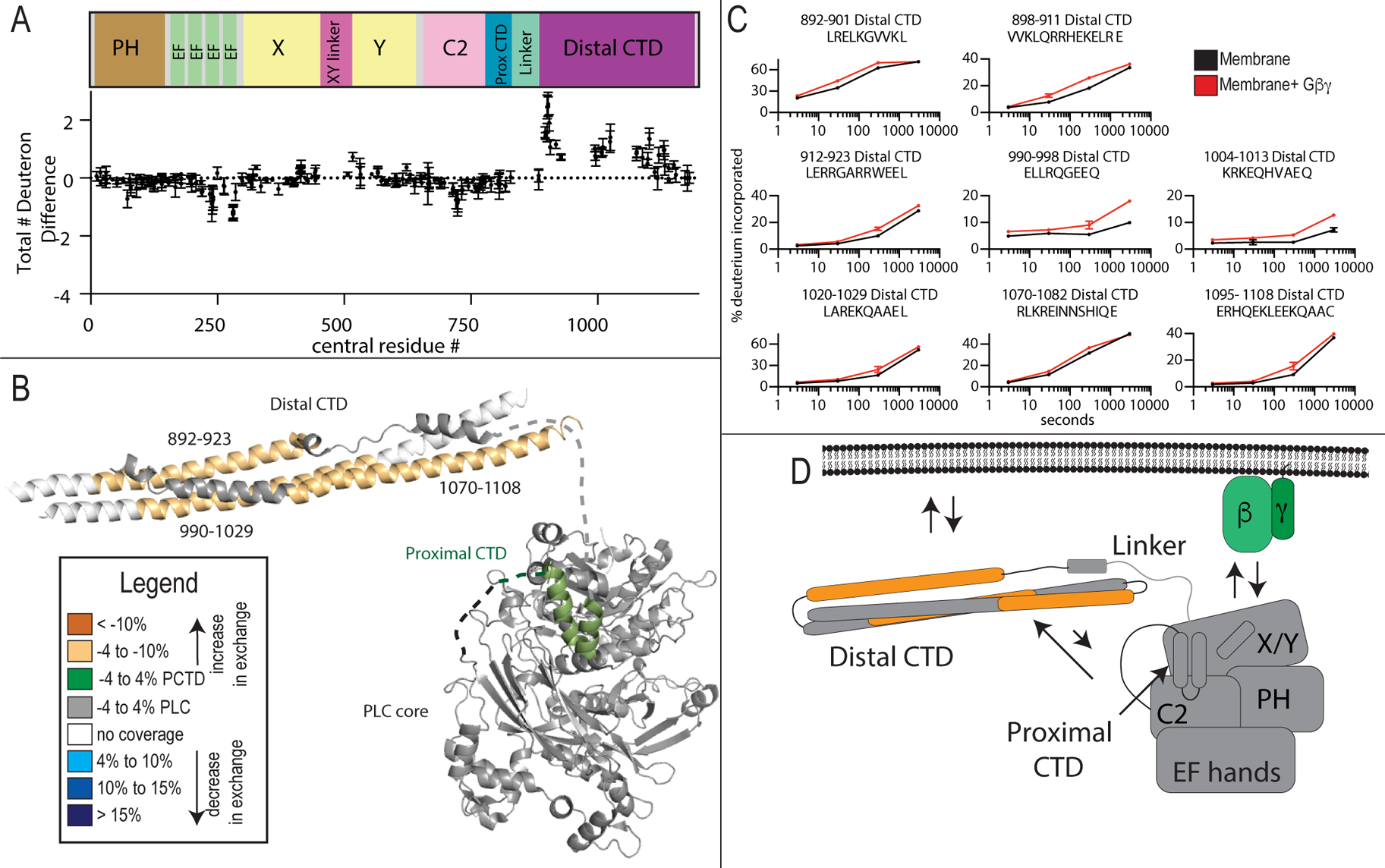
References
-
- Adjobo-Hermans MJ, Crosby KC, Putyrski M, Bhageloe A, van Weeren L, Schultz C, Goedhart J, and Gadella TW Jr. (2013). PLCβ isoforms differ in their subcellular location and their CT-domain dependent interaction with Gαq. Cell Signal 25, 255–263. - PubMed
-
- Berridge MJ (1987). Inositol tisphosphate and diacylglycerol: two interacting second messengers. Ann Rev Biochem 56, 159–193. - PubMed
-
- Bonacci TM, Ghosh M, Malik S, and Smrcka AV (2005). Regulatory interactions between the amino terminus of G-protein βγ subunits and the catalytic domain of PLCβ2. J Biol Chem 280, 10174–10181. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
