

Maternal Anti-Dengue IgG Fucosylation Predicts Susceptibility to Dengue Disease in Infants
- PMID: 32402275
- PMCID: PMC7344335
- DOI: 10.1016/j.celrep.2020.107642
Maternal Anti-Dengue IgG Fucosylation Predicts Susceptibility to Dengue Disease in Infants
Abstract
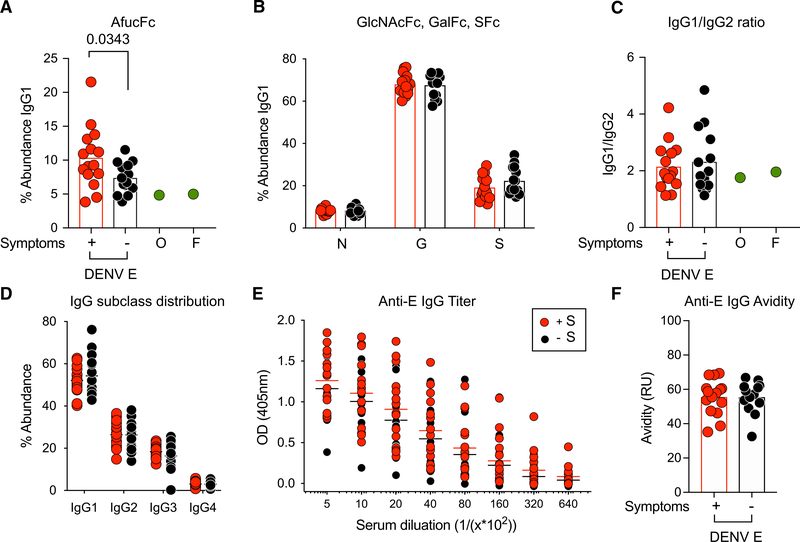
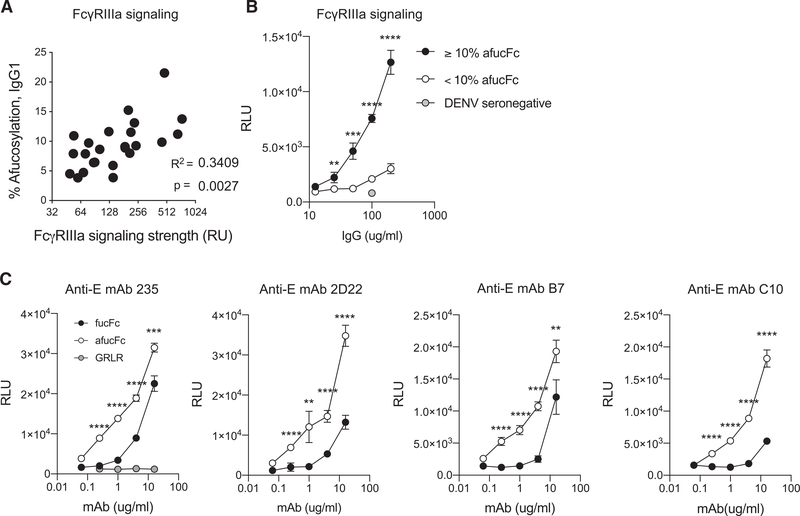
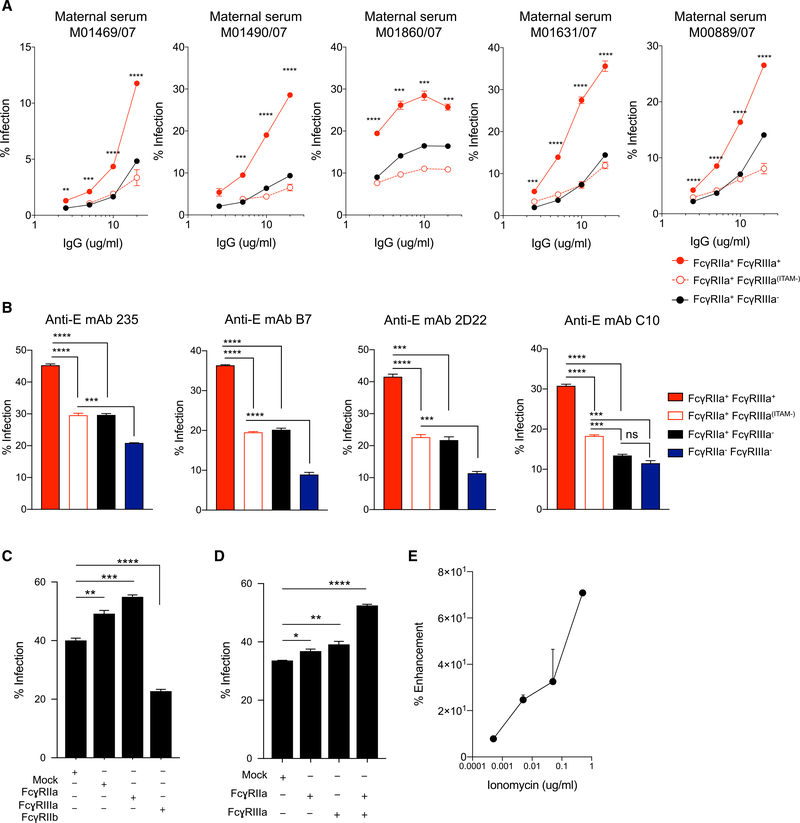
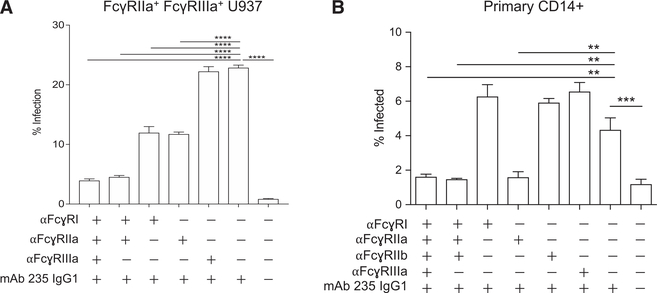
Infant mortality from dengue disease is a devastating global health burden that could be minimized with the ability to identify susceptibility for severe disease prior to infection. Although most primary infant dengue infections are asymptomatic, maternally derived anti-dengue immunoglobulin G (IgGs) present during infection can trigger progression to severe disease through antibody-dependent enhancement mechanisms. Importantly, specific characteristics of maternal IgGs that herald progression to severe infant dengue are unknown. Here, we define ≥10% afucosylation of maternal anti-dengue IgGs as a risk factor for susceptibility of infants to symptomatic dengue infections. Mechanistic experiments show that afucosylation of anti-dengue IgGs promotes FcγRIIIa signaling during infection, in turn enhancing dengue virus replication in FcγRIIIa+ monocytes. These studies identify a post-translational modification of anti-dengue IgGs that correlates with risk for symptomatic infant dengue infections and define a mechanism by which afucosylated antibodies and FcγRIIIa enhance dengue infections.
Keywords: CD16; FcγRIIIa; IgG fucosylation; antibody glycosylation; infant dengue; severe dengue.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
References
-
- Alkan Ozdemir S, Ozer EA, Kose S, Ilhan O, Ozturk C, and Sutcuoglu S (2016). Reference values of serum IgG and IgM levels in preterm and term new-borns. J. Matern. Fetal Neonatal Med. 29, 972–976. - PubMed
-
- Anderson KB, Gibbons RV, Cummings DA, Nisalak A, Green S, Libraty DH, Jarman RG, Srikiatkhachorn A, Mammen MP, Darunee B, et al. (2014). A shorter time interval between first and second dengue infections is associated with protection from clinical illness in a school-based cohort in Thailand. J. Infect. Dis. 209, 360–368. - PMC - PubMed
-
- Aramburu J, Yaffe MB, López-Rodríguez C, Cantley LC, Hogan PG, and Rao A (1999). Affinity-driven peptide selection of an NFAT inhibitor more selective than cyclosporin A. Science 285, 2129–2133. - PubMed
-
- Aye KS, Charngkaew K, Win N, Wai KZ, Moe K, Punyadee N, Thiemmeca S, Suttitheptumrong A, Sukpanichnant S, Prida M, and Halstead SB (2014). Pathologic highlights of dengue hemorrhagic fever in 13 autopsy cases from Myanmar. Hum. Pathol. 45, 1221–1233. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
