

Therapeutic potential of Nitazoxanide against Newcastle disease virus: A possible modulation of host cytokines
- PMID: 32403005
- PMCID: PMC7196422
- DOI: 10.1016/j.cyto.2020.155115
Therapeutic potential of Nitazoxanide against Newcastle disease virus: A possible modulation of host cytokines
Abstract
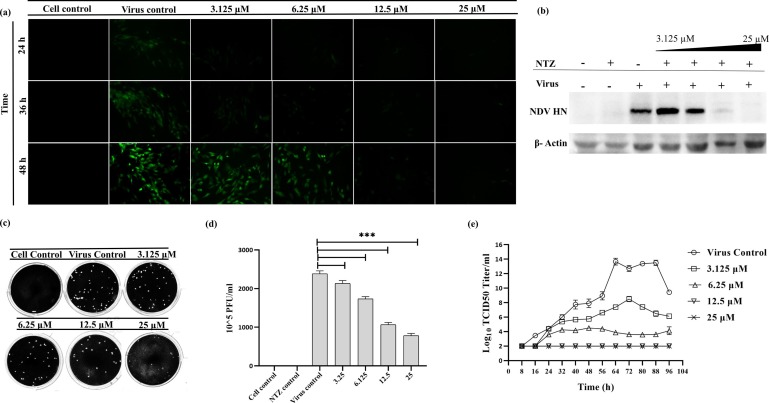
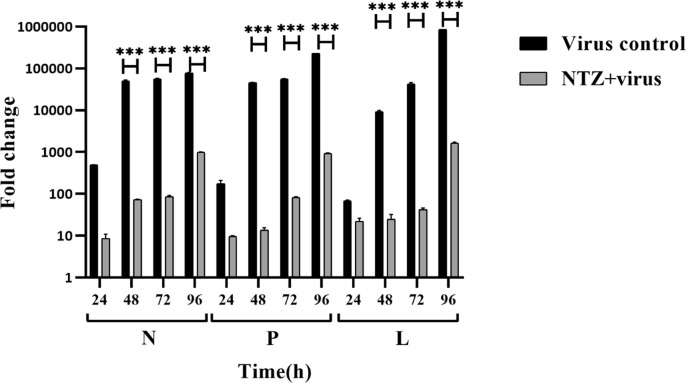
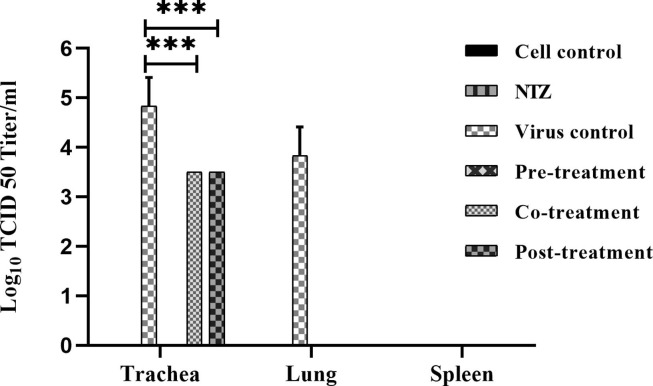
Newcastle disease (ND) is prevalent among the domesticated and the wild birds and is caused by the avian paramyxovirus serotype-I (APMV-I). It is commonly known to affect chicken, pheasant, ostrich, pigeon and waterfowl. Depending on the virulence, the velogenic NDV strains cause severe respiratory and nervous disorders with a high mortality rate. The live and killed vaccines are available for the prevention of infection in the market, but the drug for the treatment is not available. Nitazoxanide (NTZ), a member of thiazolides, is an antiparasitic drug. In the present study, the effect of NTZ on the NDV replication was explored. The experiments were conducted in chicken fibroblast cells (DF-1), PBMC, embryonated chicken eggs, and two-week old chickens. The inhibition of the NDV was observed upon post-treatment of NTZ at a concentration of ~12.5 μM. Cytokine profiling of the DF-1, PBMC, and chicken embryonic tissue treated with NTZ revealed significant upregulation in all the cytokines studied except for IL-1β in DF-1 cells. It is plausible that NTZ is involved in causing immune-modulatory effects in poultry. NTZ treatment in two weeks old chicken showed significant reduction in NDV replication in trachea, and lungs, respectively, at 72 h post-infection. Encouraging results from the present study warrants repurposing NTZ as a drug for the treatment of viral infection in poultry. It will also pave the way towards understanding of similar effect against other animal pathogens.
Keywords: Avian paramyxovirus; Cytokines; In vitro; Nitazoxanide; Repurposing.
Copyright © 2020 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures




References
-
- R.A. Lamb, G.D. Parks, Paramyxoviridae: The Viruses and their Replication, 5th ed., Lippincott, Williams, and Wilkins, 2007.
-
- Czegledi A., Ujvari D., Somogyi E., Wehmann E., Werner O., Lomniczi B. Third genome size category of avian paramyxovirus serotype 1 (Newcastle disease virus) and evolutionary implications. Virus Res. 2006;120(1–2):36–48. - PubMed
-
- de Leeuw O., Peeters B. Complete nucleotide sequence of Newcastle disease virus: evidence for the existence of a new genus within the subfamily Paramyxovirinae. J. Gen. Virol. 1999;80(Pt 1):131–136. - PubMed
-
- Huang Y., Wan H.Q., Liu H.Q., Wu Y.T., Liu X.F. Genomic sequence of an isolate of Newcastle disease virus isolated from an outbreak in geese: a novel six nucleotide insertion in the non-coding region of the nucleoprotein gene. Brief Rep. Arch. Virol. 2004;149(7):1445–1457. - PubMed
-
- Krishnamurthy S., Samal S.K. Nucleotide sequences of the trailer, nucleocapsid protein gene and intergenic regions of Newcastle disease virus strain Beaudette C and completion of the entire genome sequence. J. Gen. Virol. 1998;79(Pt 10):2419–2424. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
