

The many faces of Polycomb regulation by RNA
- PMID: 32403014
- PMCID: PMC7653676
- DOI: 10.1016/j.gde.2020.02.023
The many faces of Polycomb regulation by RNA
Abstract
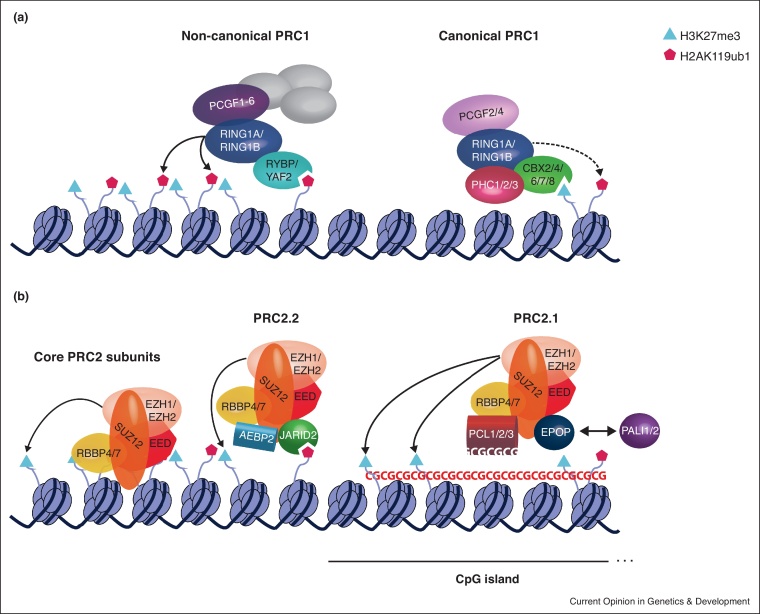
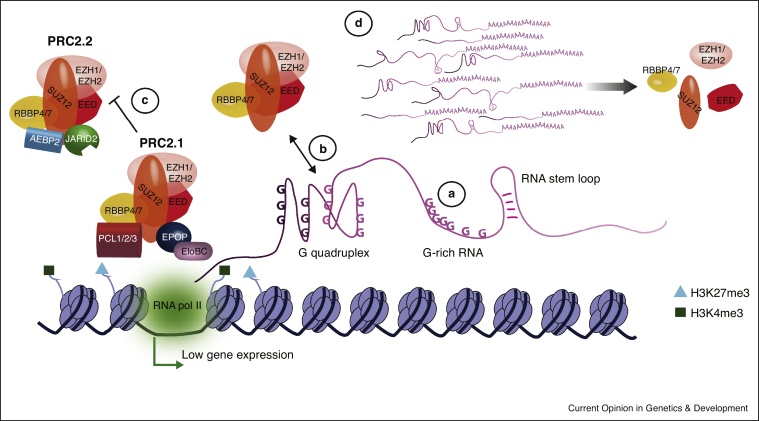
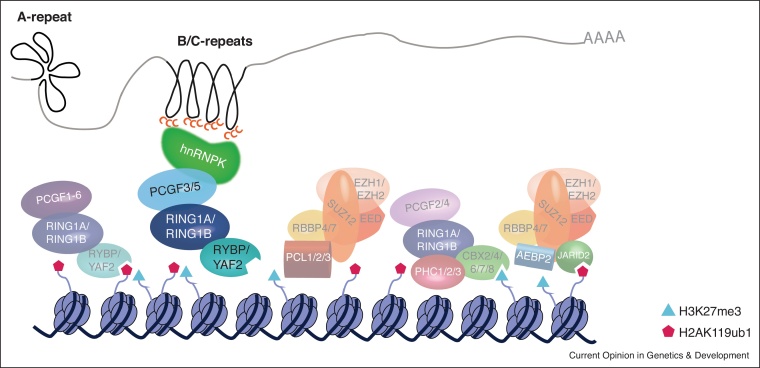
Many intricate pathways contribute to the timely control of gene expression during development. Polycomb repressive complexes (PRC1 and PRC2) and long non-coding RNAs (lncRNAs) are players associated with gene repression in various developmental processes such as X chromosome inactivation (XCI) and genomic imprinting. Historically, lncRNAs were proposed to directly recruit PRC2. However, recent evidence suggests that promiscuous interactions between PRC2 and RNA fine-tune the function of the complex through a multiplicity of mechanisms. A PRC2-recruitment model was definitively overturned in the paradigm of XCI by Xist RNA, being replaced by a novel mechanism which puts PRC1 in the spotlight. This review focuses on these recent advances in understanding the interplay between RNA and Polycomb complexes for gene expression control.
Copyright © 2020 The Author(s). Published by Elsevier Ltd.. All rights reserved.
Figures
References
-
- Kennison J.A. Methods in Enzymology. Elsevier; 2003. Introduction to Trx-G and Pc-G genes; pp. 61–70.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
