

Downregulation of parvalbumin expression in the prefrontal cortex during adolescence causes enduring prefrontal disinhibition in adulthood
- PMID: 32403119
- PMCID: PMC7360578
- DOI: 10.1038/s41386-020-0709-9
Downregulation of parvalbumin expression in the prefrontal cortex during adolescence causes enduring prefrontal disinhibition in adulthood
Abstract
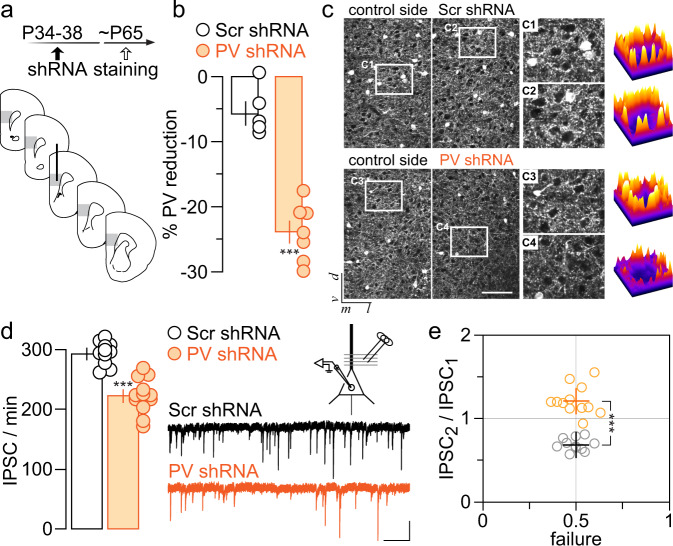
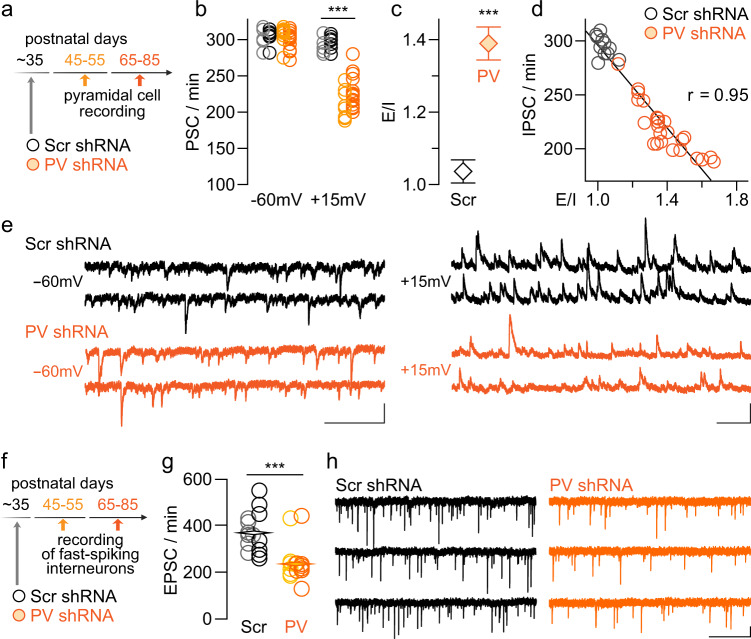
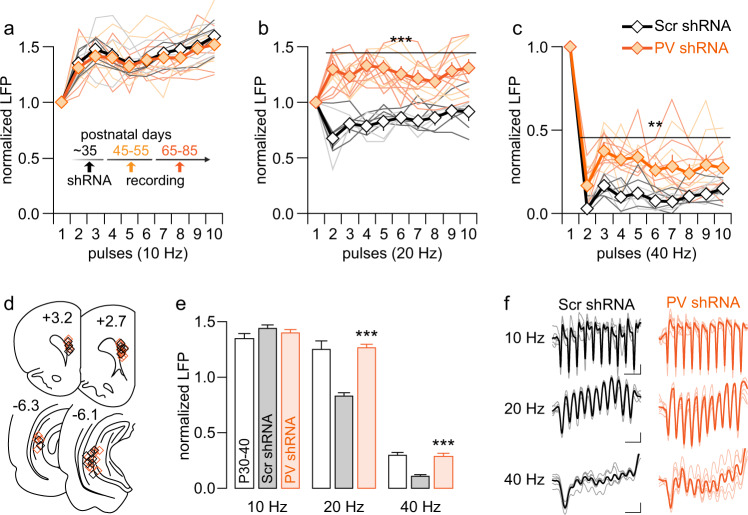
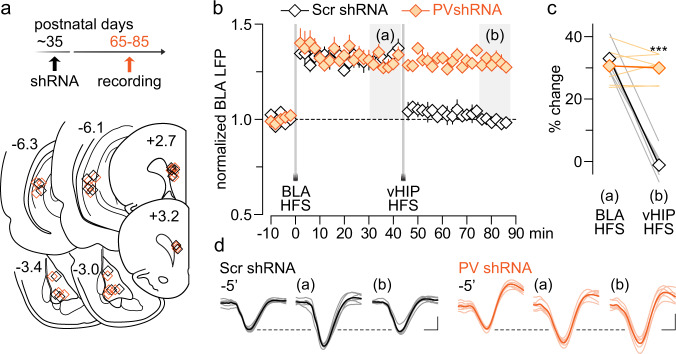
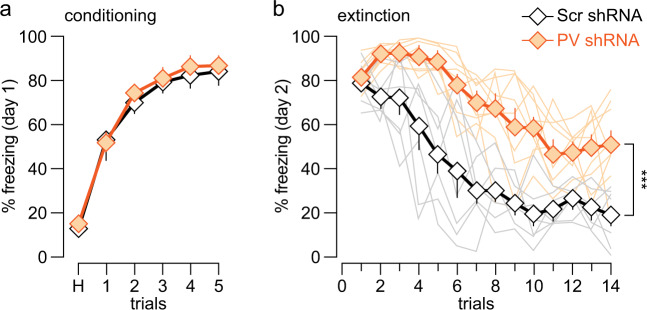
The expression of the calcium binding protein parvalbumin (PV) has been observed in several cortical regions during development in a temporal pattern consistent with increased afferent-dependent activity. In the prefrontal cortex (PFC), PV expression appears last and continues to substantially increase throughout adolescence, yet the significance of this increase remains unclear. Because of the expression of PV in fast-spiking GABAergic interneurons, we hypothesized that PV upregulation during adolescence is necessary to sustain the increase in GABAergic activity observed in the PFC during this period. To test this hypothesis, we utilized an RNAi strategy to directly downregulate PV levels in the PFC during adolescence and examined its impact on prefrontal GABAergic function, plasticity, and associated behaviors during adulthood. The data indicate that a mere 25% reduction of adult PV levels in the PFC was sufficient to reduce local GABAergic transmission onto pyramidal neurons, disrupt prefrontal excitatory-inhibitory balance, and alter processing of afferent information from the ventral hippocampus. Accordingly, these animals displayed an impairment in the level of extinction learning of a trace fear conditioning response, a behavioral paradigm that requires intact PFC-ventral hippocampus connectivity. These results indicate the PV upregulation observed in the PFC during adolescence is necessary for refinement of prefrontal GABAergic function, the absence of which results in immature afferent processing and a hypofunctional state. Importantly, these results suggest there is a critical window of plasticity during which PV upregulation supports the acquisition of mature GABAergic phenotype necessary to sustain adult PFC functions.
Figures
Similar articles
-
MK-801 Exposure during Adolescence Elicits Enduring Disruption of Prefrontal E-I Balance and Its Control of Fear Extinction Behavior.J Neurosci. 2020 Jun 17;40(25):4881-4887. doi: 10.1523/JNEUROSCI.0581-20.2020. Epub 2020 May 19. J Neurosci. 2020. PMID: 32430298 Free PMC article.
-
Differential regulation of parvalbumin and calretinin interneurons in the prefrontal cortex during adolescence.Brain Struct Funct. 2014 Jan;219(1):395-406. doi: 10.1007/s00429-013-0508-8. Epub 2013 Feb 12. Brain Struct Funct. 2014. PMID: 23400698 Free PMC article.
-
Changes in the Prefrontal Glutamatergic and Parvalbumin Systems of Mice Exposed to Unpredictable Chronic Stress.Mol Neurobiol. 2018 Mar;55(3):2591-2602. doi: 10.1007/s12035-017-0528-0. Epub 2017 Apr 18. Mol Neurobiol. 2018. PMID: 28421533
-
Prefrontal GABAergic Interneurons Gate Long-Range Afferents to Regulate Prefrontal Cortex-Associated Complex Behaviors.Front Neural Circuits. 2021 Jul 12;15:716408. doi: 10.3389/fncir.2021.716408. eCollection 2021. Front Neural Circuits. 2021. PMID: 34322002 Free PMC article. Review.
-
Prefrontal and Hippocampal Parvalbumin Interneurons in Animal Models for Schizophrenia: A Systematic Review and Meta-analysis.Schizophr Bull. 2024 Jan 1;50(1):210-223. doi: 10.1093/schbul/sbad123. Schizophr Bull. 2024. PMID: 37584417 Free PMC article.
Cited by
-
A transient developmental increase in prefrontal activity alters network maturation and causes cognitive dysfunction in adult mice.Neuron. 2021 Apr 21;109(8):1350-1364.e6. doi: 10.1016/j.neuron.2021.02.011. Epub 2021 Mar 5. Neuron. 2021. PMID: 33675685 Free PMC article.
-
Development of the rodent prefrontal cortex: circuit formation, plasticity, and impacts of early life stress.Front Neural Circuits. 2025 Mar 26;19:1568610. doi: 10.3389/fncir.2025.1568610. eCollection 2025. Front Neural Circuits. 2025. PMID: 40206866 Free PMC article. Review.
-
Gene-environmental regulation of the postnatal post-mitotic neuronal maturation.Trends Genet. 2024 Jun;40(6):480-494. doi: 10.1016/j.tig.2024.03.006. Epub 2024 Apr 23. Trends Genet. 2024. PMID: 38658255 Free PMC article. Review.
-
GABA System Modifications During Periods of Hormonal Flux Across the Female Lifespan.Front Behav Neurosci. 2022 Jun 16;16:802530. doi: 10.3389/fnbeh.2022.802530. eCollection 2022. Front Behav Neurosci. 2022. PMID: 35783228 Free PMC article. Review.
-
Uniform volumetric single-cell processing for organ-scale molecular phenotyping.Nat Biotechnol. 2025 Jan 24. doi: 10.1038/s41587-024-02533-4. Online ahead of print. Nat Biotechnol. 2025. PMID: 39856430
References
-
- Giedd JN, Blumenthal J, Jeffries NO, Castellanos FX, Liu H, Zijdenbos A, et al. Brain development during childhood and adolescence: a longitudinal MRI study. Nat Neurosci. 1999;2:861–3. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
