

TENSCell: Imaging of Stretch-Activated Cells Reveals Divergent Nuclear Behavior and Tension
- PMID: 32407683
- PMCID: PMC7264846
- DOI: 10.1016/j.bpj.2020.03.035
TENSCell: Imaging of Stretch-Activated Cells Reveals Divergent Nuclear Behavior and Tension
Abstract
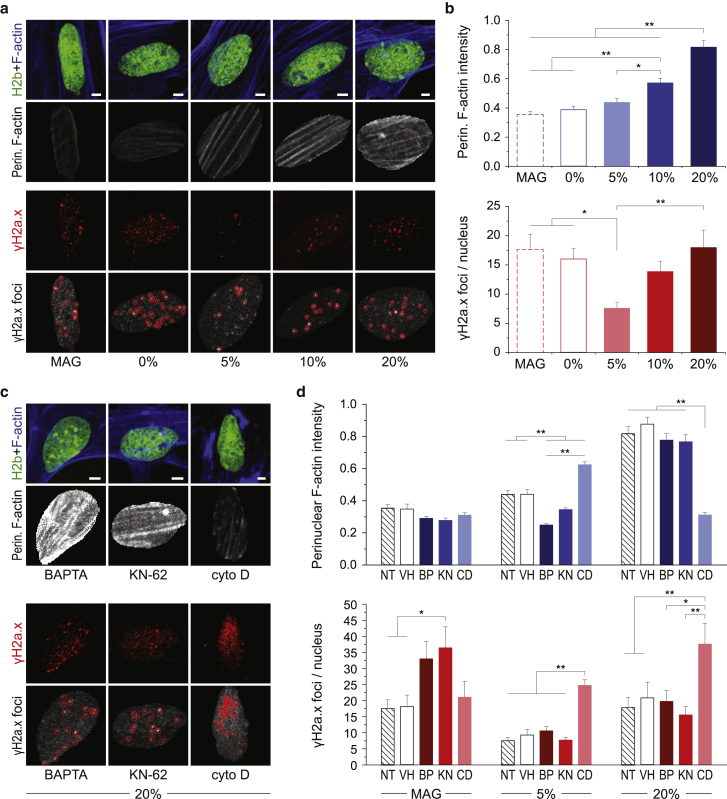
Mechanisms of cellular and nuclear mechanosensation are unclear, partially because of a lack of methods that can reveal dynamic processes. Here, we present a new concept for a low-cost, three-dimensionally printed device that enables high-magnification imaging of cells during stretch. We observed that nuclei of mouse embryonic skin fibroblasts underwent rapid (within minutes) and divergent responses, characterized by nuclear area expansion during 5% strain but nuclear area shrinkage during 20% strain. Only responses to low strain were dependent on calcium signaling, whereas actin inhibition abrogated all nuclear responses and increased nuclear strain transfer and DNA damage. Imaging of actin dynamics during stretch revealed similar divergent trends, with F-actin shifting away from (5% strain) or toward (20% strain) the nuclear periphery. Our findings emphasize the importance of simultaneous stimulation and data acquisition to capture mechanosensitive responses and suggest that mechanical confinement of nuclei through actin may be a protective mechanism during high mechanical stretch or loading.
Copyright © 2020 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures





References
-
- Engler A.J., Sen S., Discher D.E. Matrix elasticity directs stem cell lineage specification. Cell. 2006;126:677–689. - PubMed
-
- Hemphill M.A., Dauth S., Parker K.K. Traumatic brain injury and the neuronal microenvironment: a potential role for neuropathological mechanotransduction. Neuron. 2015;85:1177–1192. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
