

ERα-related chromothripsis enhances concordant gene transcription on chromosome 17q11.1-q24.1 in luminal breast cancer
- PMID: 32408897
- PMCID: PMC7222439
- DOI: 10.1186/s12920-020-0729-7
ERα-related chromothripsis enhances concordant gene transcription on chromosome 17q11.1-q24.1 in luminal breast cancer
Abstract
Background: Chromothripsis is an event of genomic instability leading to complex chromosomal alterations in cancer. Frequent long-range chromatin interactions between transcription factors (TFs) and targets may promote extensive translocations and copy-number alterations in proximal contact regions through inappropriate DNA stitching. Although studies have proposed models to explain the initiation of chromothripsis, few discussed how TFs influence this process for tumor progression.
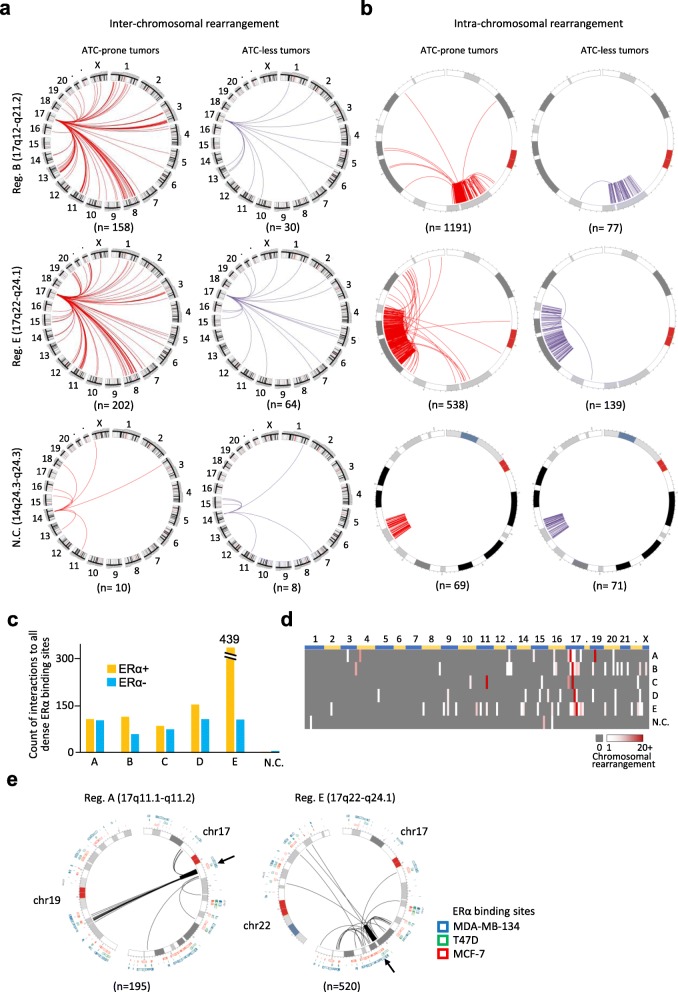
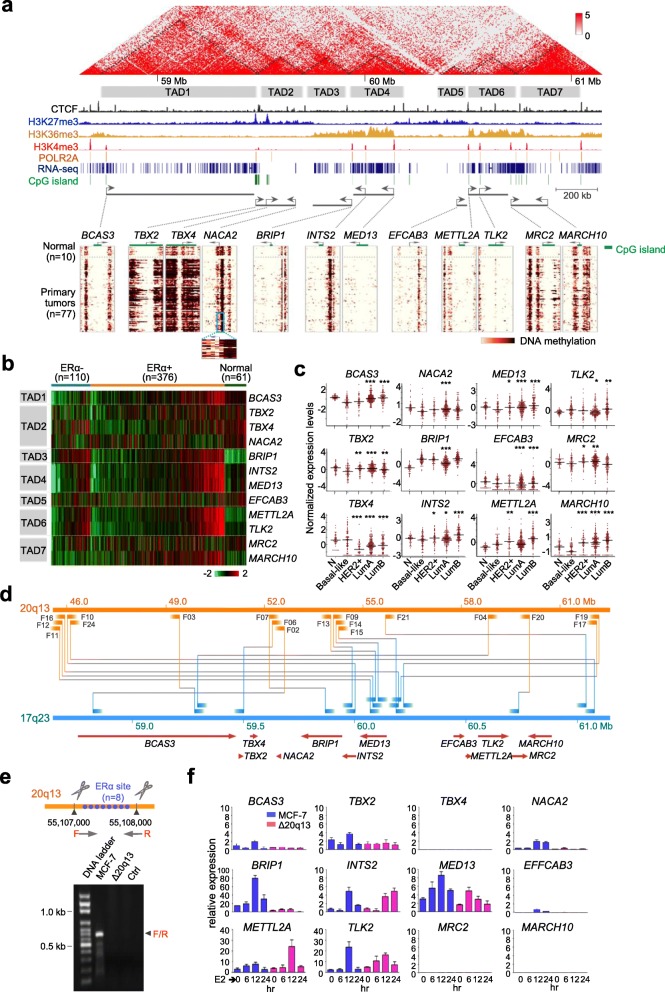
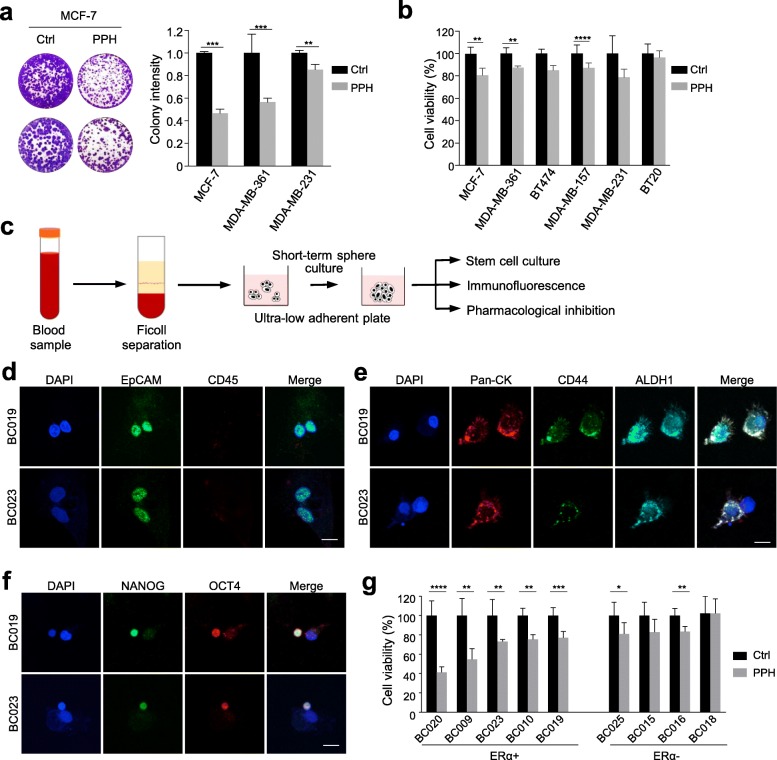
Methods: This study focused on genomic alterations in amplification associated regions within chromosome 17. Inter-/intra-chromosomal rearrangements were analyzed using whole genome sequencing data of breast tumors in the Cancer Genome Atlas (TCGA) cohort. Common ERα binding sites were defined based on MCF-7, T47D, and MDA-MB-134 breast cancer cell lines using univariate K-means clustering methods. Nanopore sequencing technology was applied to validate frequent rearrangements detected between ATC loci on 17q23 and an ERα hub on 20q13. The efficacy of pharmacological inhibition of a potentially druggable target gene on 17q23 was evaluated using breast cancer cell lines and patient-derived circulating breast tumor cells.
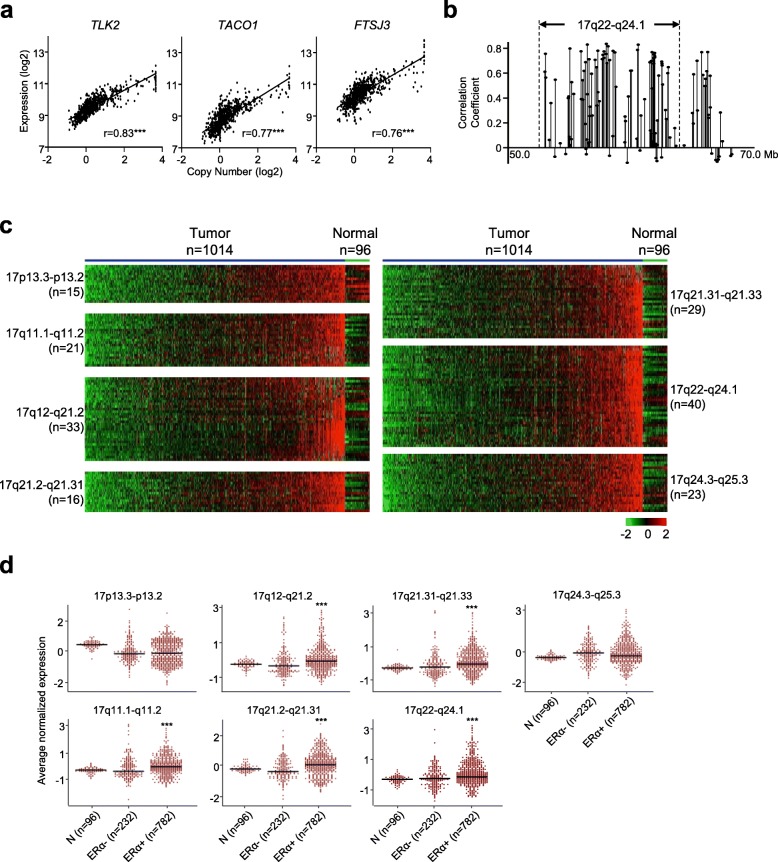
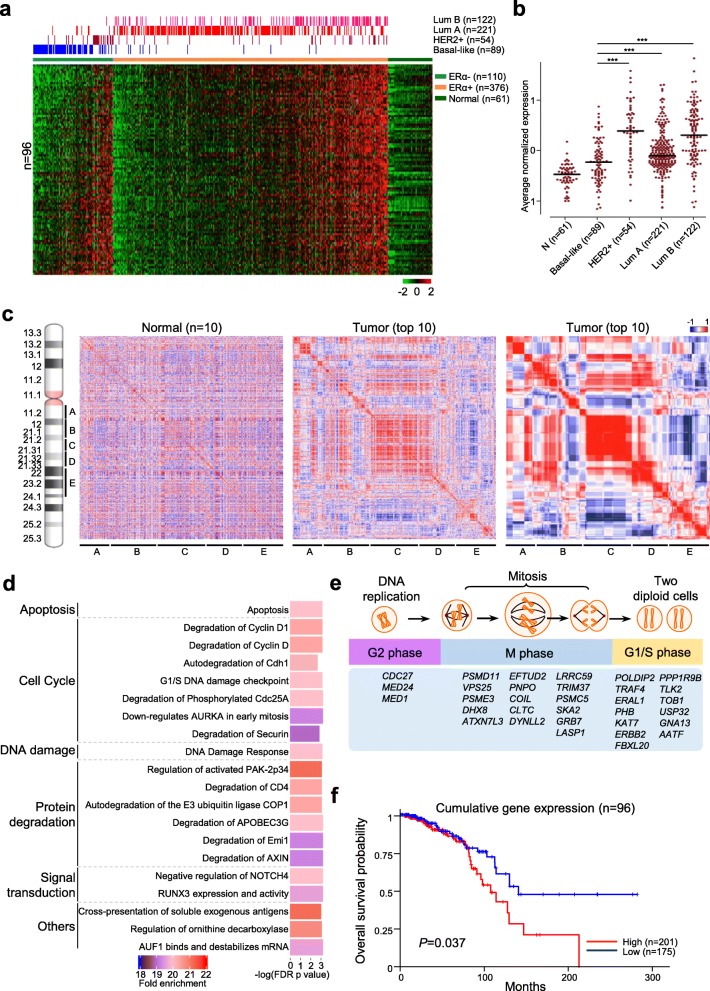
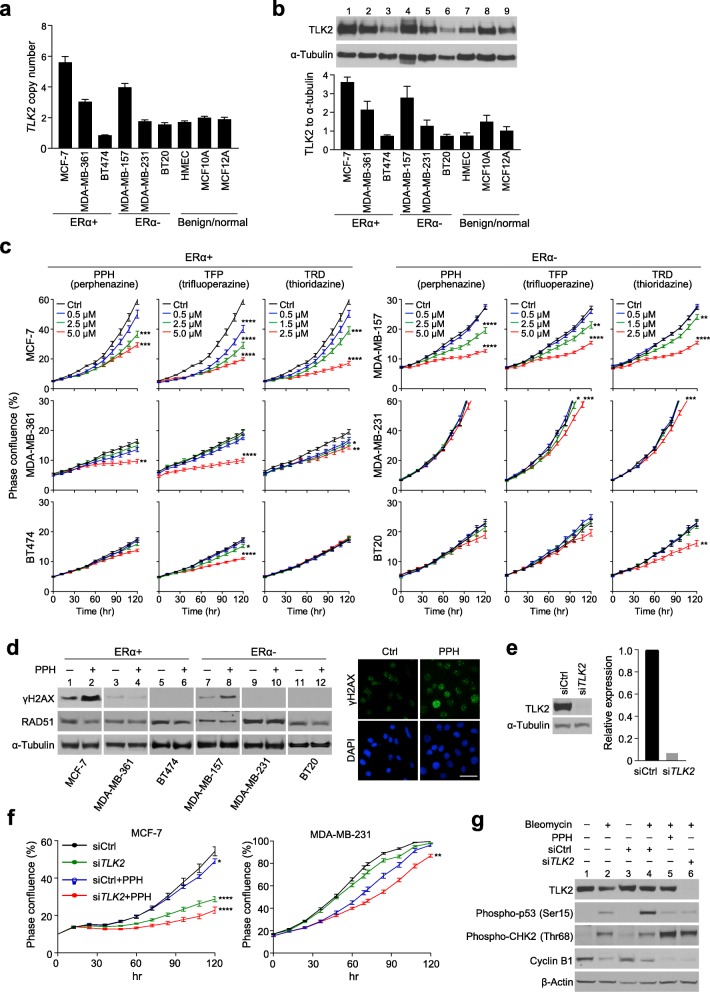
Results: There are five adjoining regions from 17q11.1 to 17q24.1 being hotspots of chromothripsis. Inter-/intra-chromosomal rearrangements of these regions occurred more frequently in ERα-positive tumors than in ERα-negative tumors. In addition, the locations of the rearrangements were often mapped within or close to dense ERα binding sites localized on these five 17q regions or other chromosomes. This chromothriptic event was linked to concordant upregulation of 96 loci that predominantly regulate cell-cycle machineries in advanced luminal tumors. Genome-editing analysis confirmed that an ERα hub localized on 20q13 coordinately regulates a subset of these loci localized on 17q23 through long-range chromosome interactions. One of these loci, Tousled Like Kinase 2 (TLK2) known to participate in DNA damage checkpoint control, is an actionable target using phenothiazine antipsychotics (PTZs). The antiproliferative effect of PTZs was prominent in high TLK2-expressing cells, compared to low expressing cells.
Conclusion: This study demonstrates a new approach for identifying tumorigenic drivers from genomic regions highly susceptible to ERα-related chromothripsis. We found a group of luminal breast tumors displaying 17q-related chromothripsis for which antipsychotics can be repurposed as treatment adjuncts.
Keywords: Chromosomal rearrangement; Chromothripsis; Concordant transcription; Druggable target; ERα; Luminal subtype breast cancer; Nanopore sequencing.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
