

De novo protein design enables the precise induction of RSV-neutralizing antibodies
- PMID: 32409444
- PMCID: PMC7391827
- DOI: 10.1126/science.aay5051
De novo protein design enables the precise induction of RSV-neutralizing antibodies
Abstract
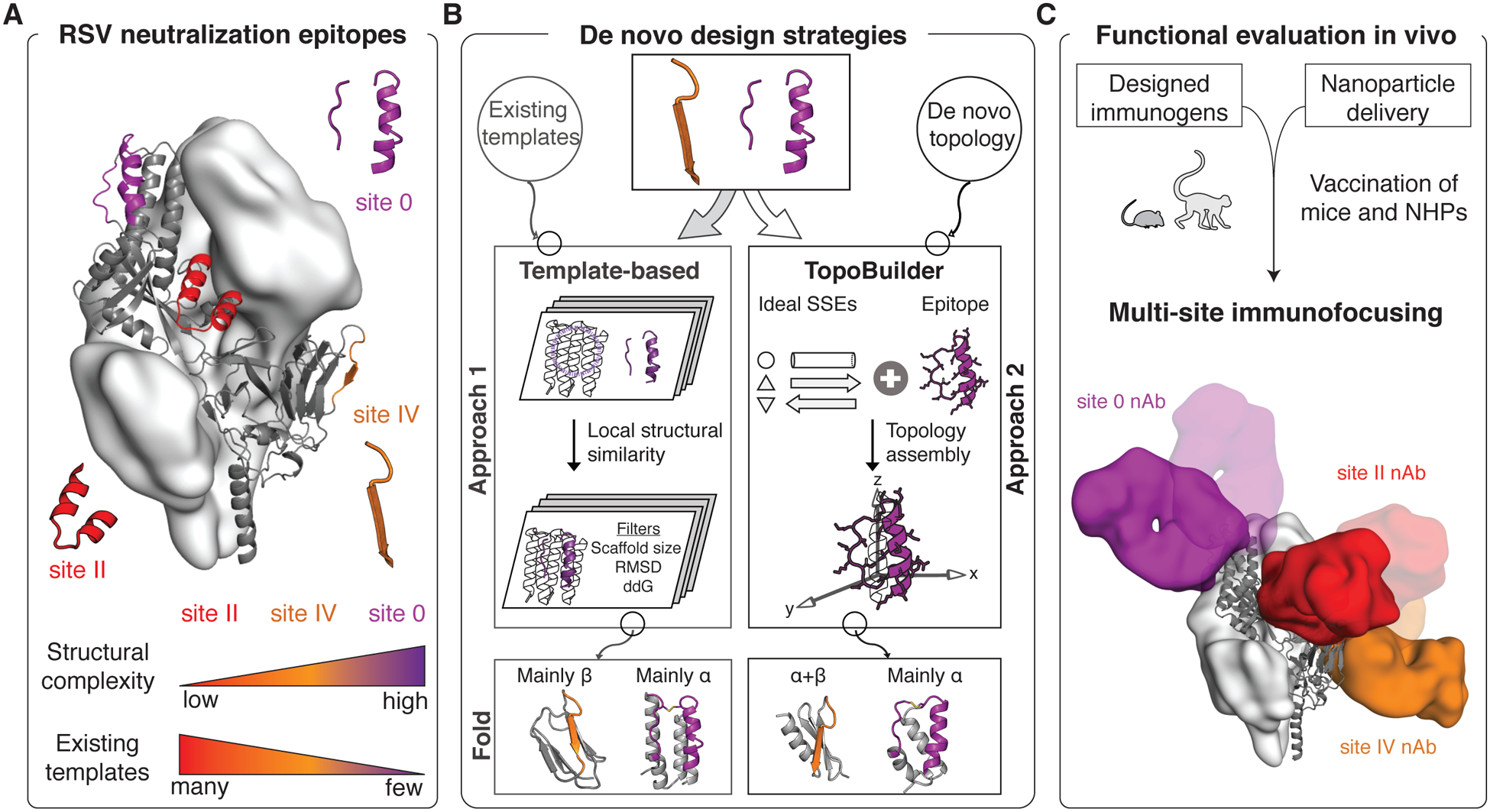
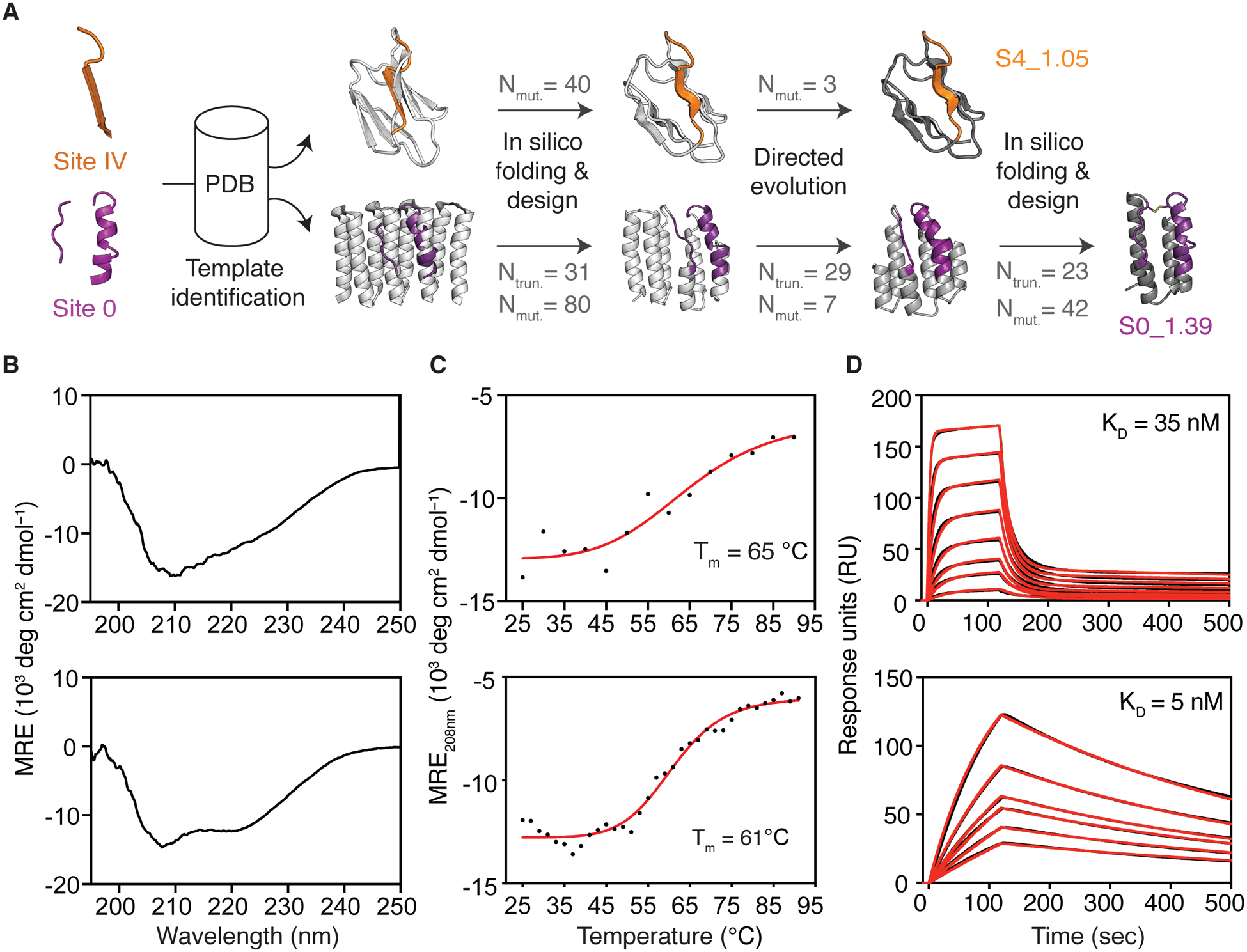
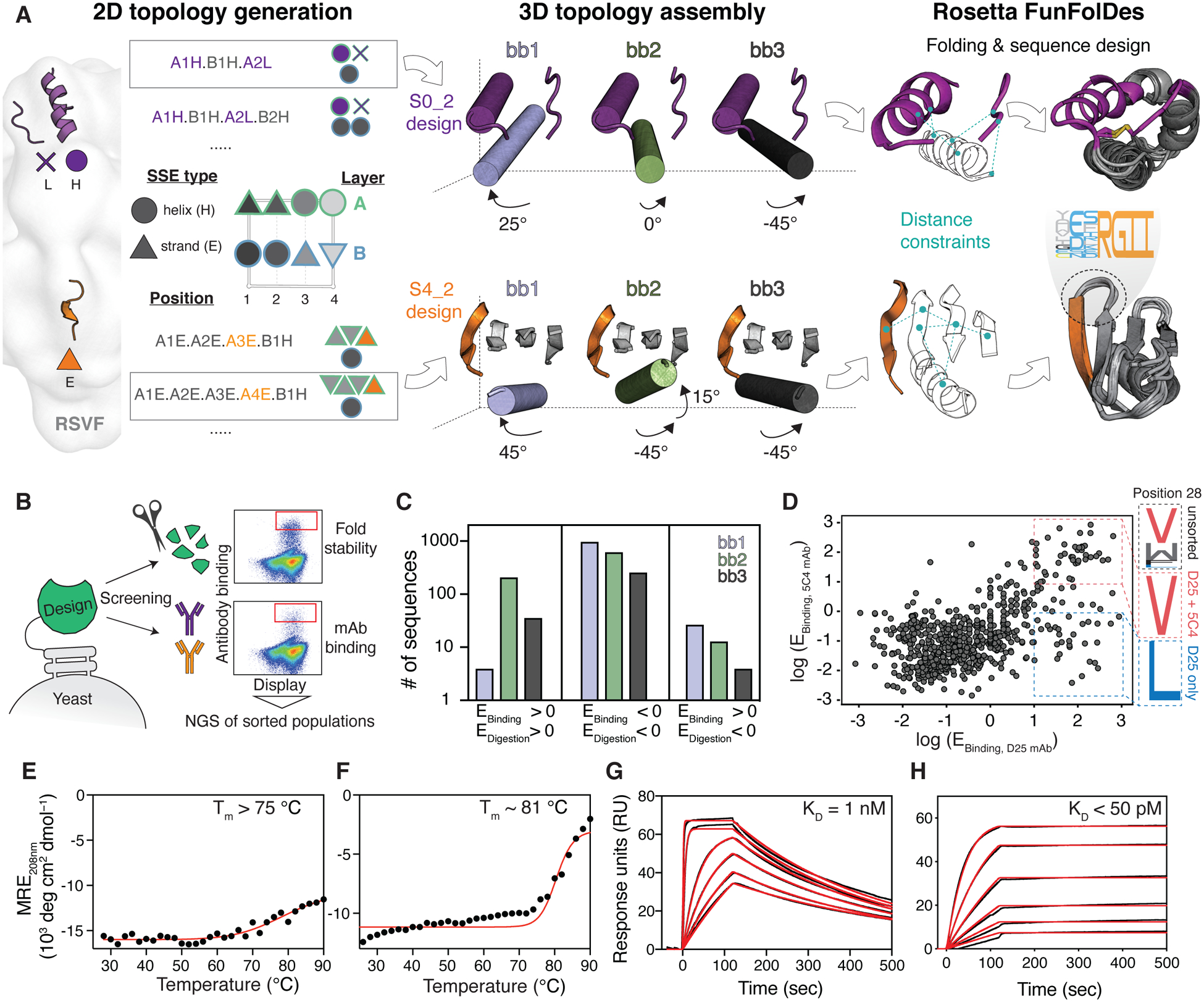
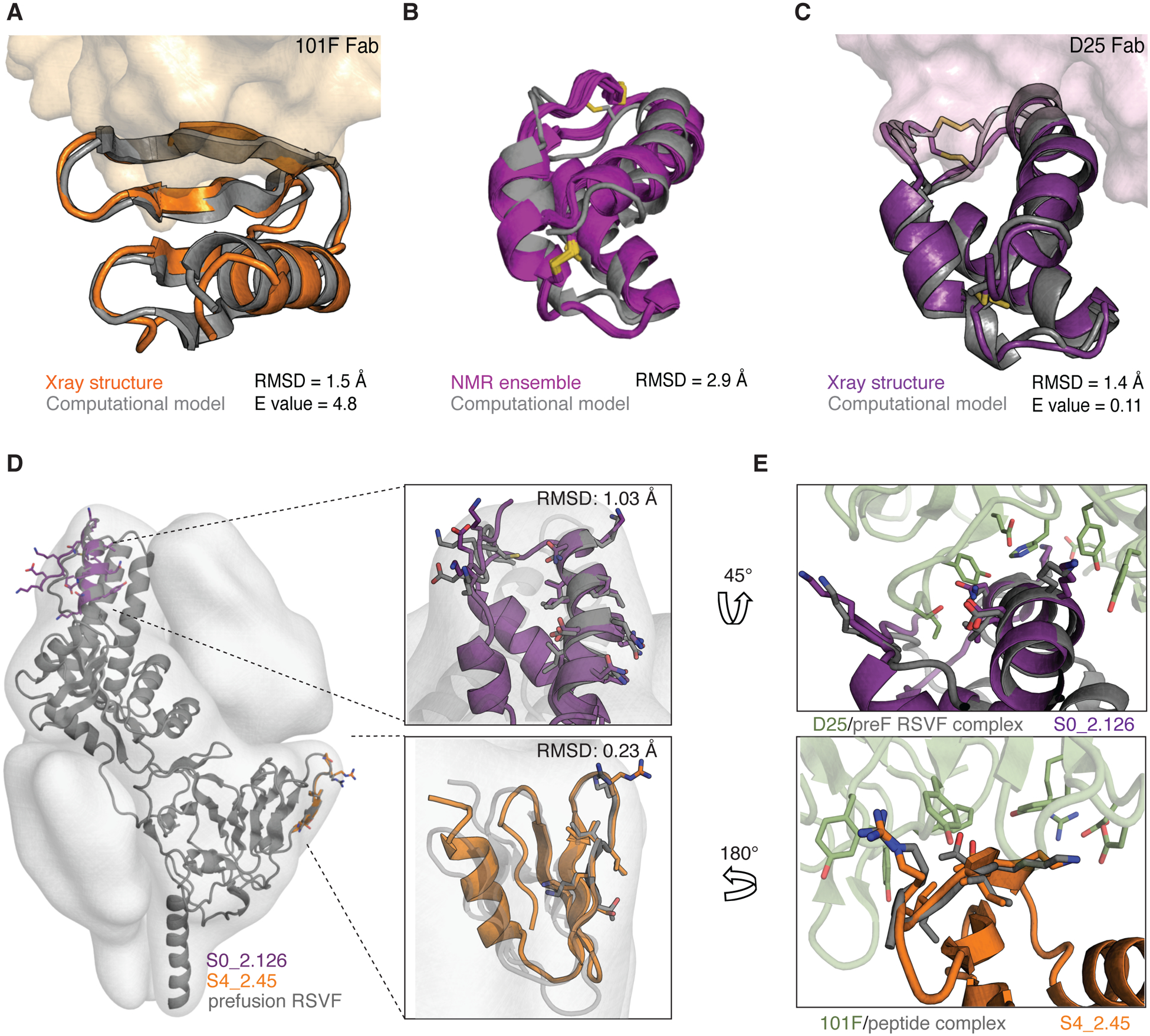
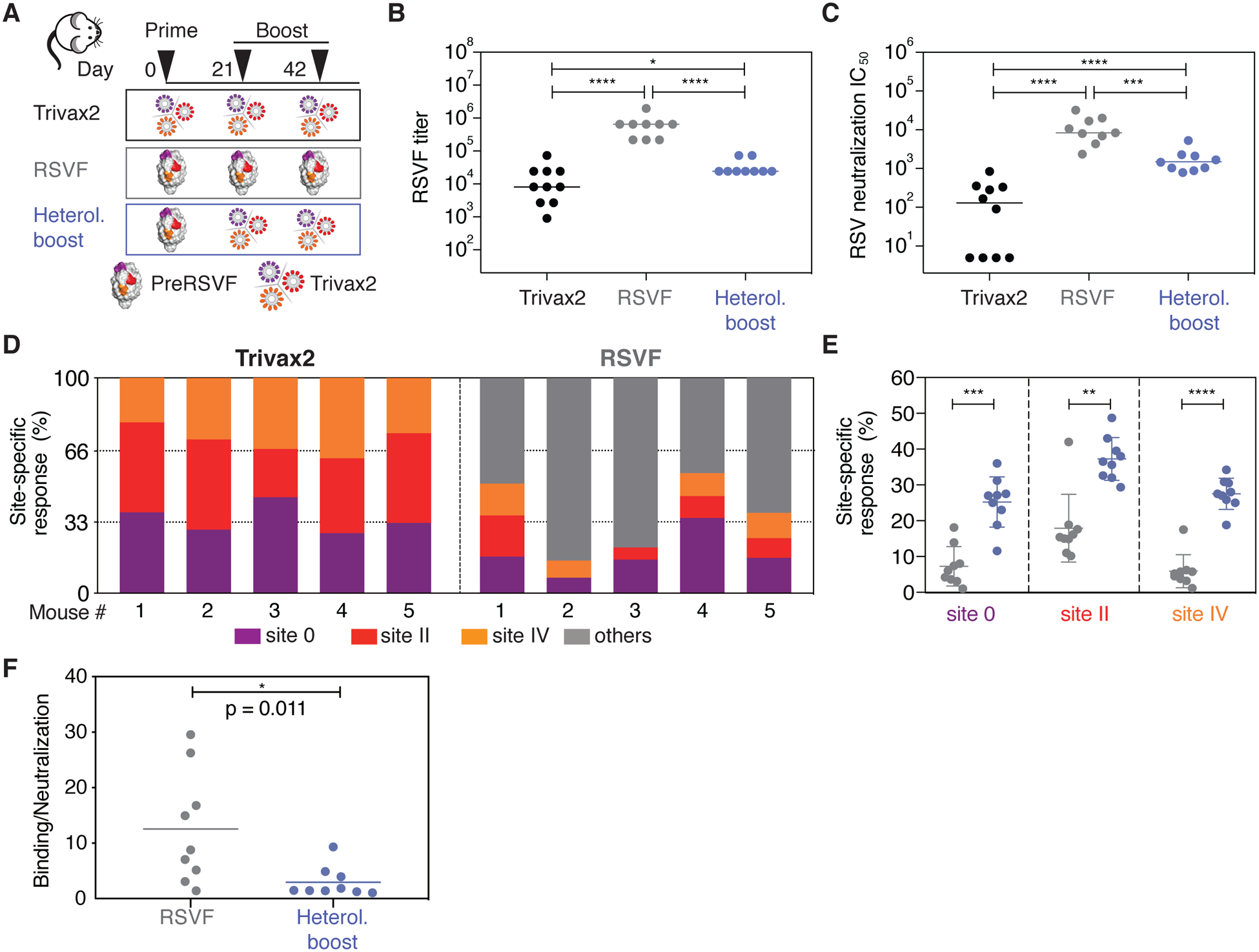
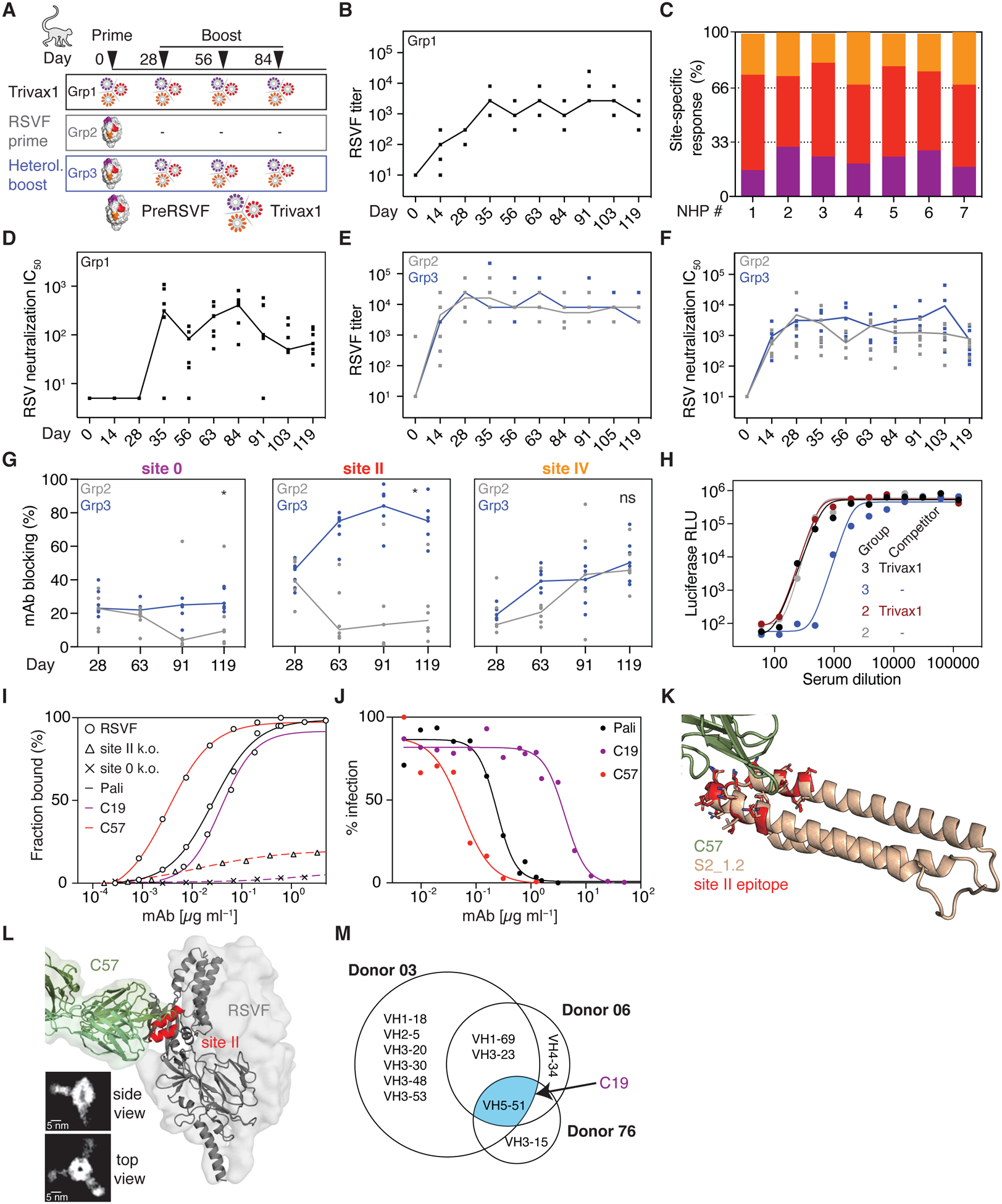
De novo protein design has been successful in expanding the natural protein repertoire. However, most de novo proteins lack biological function, presenting a major methodological challenge. In vaccinology, the induction of precise antibody responses remains a cornerstone for next-generation vaccines. Here, we present a protein design algorithm called TopoBuilder, with which we engineered epitope-focused immunogens displaying complex structural motifs. In both mice and nonhuman primates, cocktails of three de novo-designed immunogens induced robust neutralizing responses against the respiratory syncytial virus. Furthermore, the immunogens refocused preexisting antibody responses toward defined neutralization epitopes. Overall, our design approach opens the possibility of targeting specific epitopes for the development of vaccines and therapeutic antibodies and, more generally, will be applicable to the design of de novo proteins displaying complex functional motifs.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Comment in
-
Bottom-up de novo protein design.Nat Methods. 2021 Mar;18(3):233. doi: 10.1038/s41592-021-01097-4. Nat Methods. 2021. PMID: 33674794 No abstract available.
References
-
- Huang PS, Boyken SE, Baker D, The coming of age of de novo protein design. Nature 537, 320–327 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
