

Pharmacological inhibition of PRMT7 links arginine monomethylation to the cellular stress response
- PMID: 32409666
- PMCID: PMC7224190
- DOI: 10.1038/s41467-020-16271-z
Pharmacological inhibition of PRMT7 links arginine monomethylation to the cellular stress response
Erratum in
-
Author Correction: Pharmacological inhibition of PRMT7 links arginine monomethylation to the cellular stress response.Nat Commun. 2020 May 26;11(1):2683. doi: 10.1038/s41467-020-16613-x. Nat Commun. 2020. PMID: 32457299 Free PMC article.
Abstract
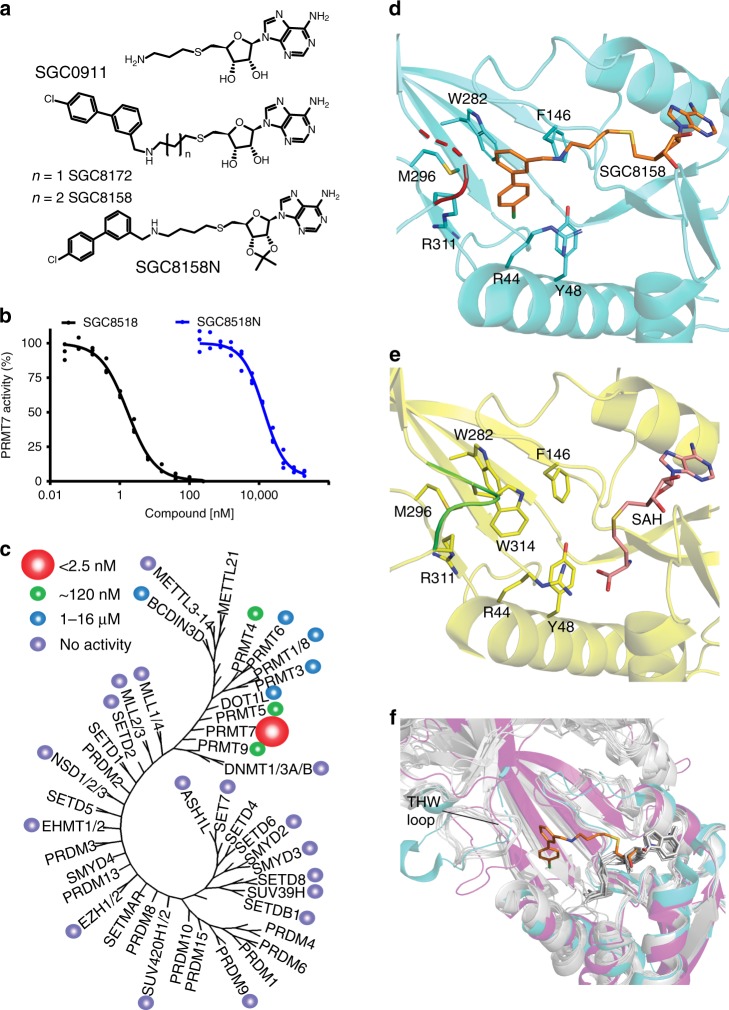
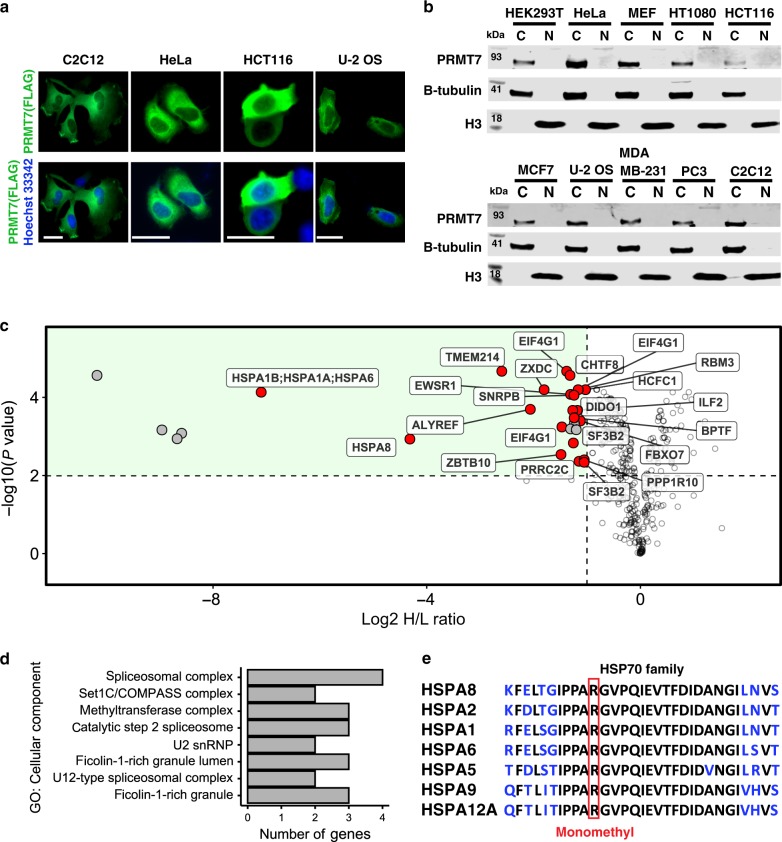
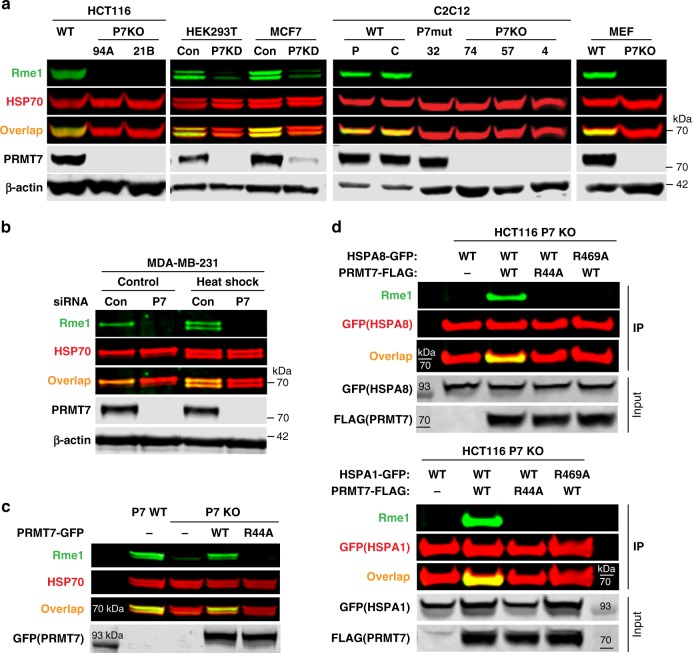
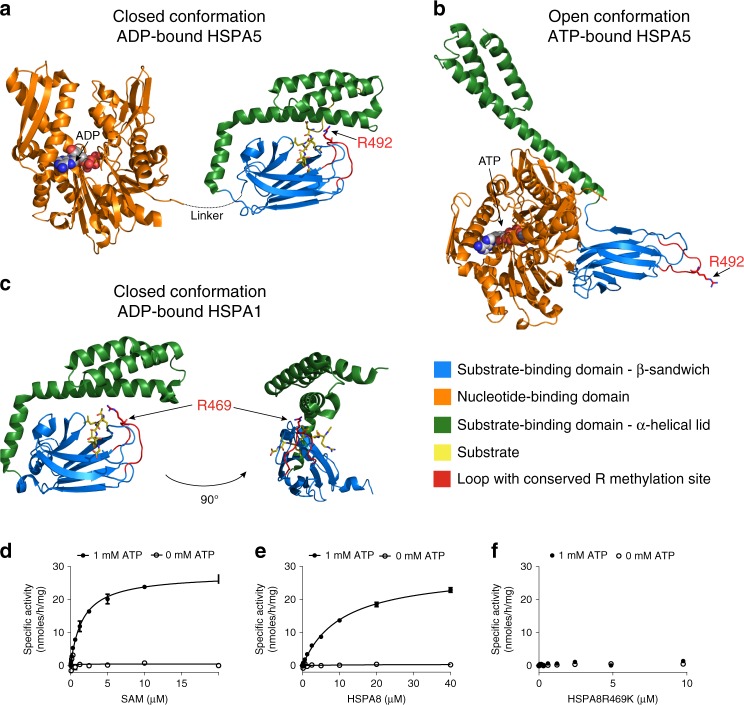
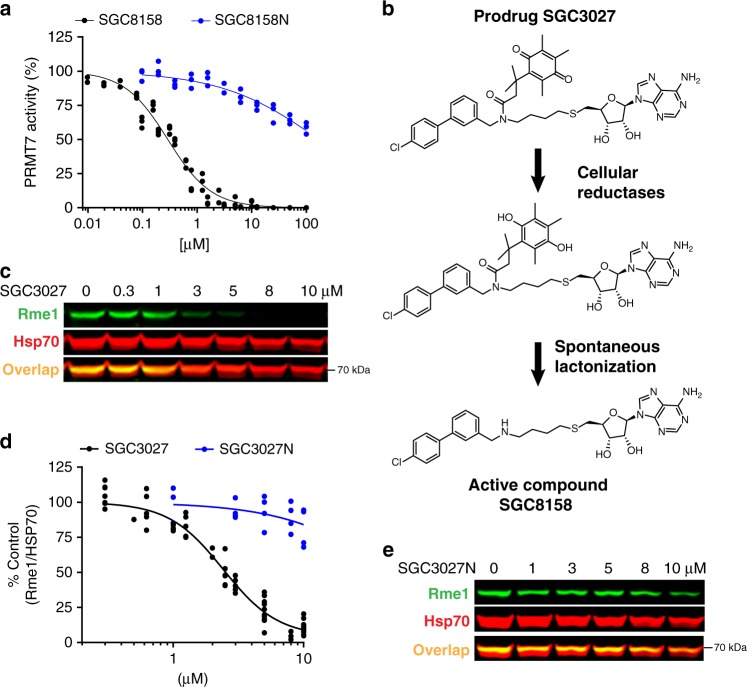
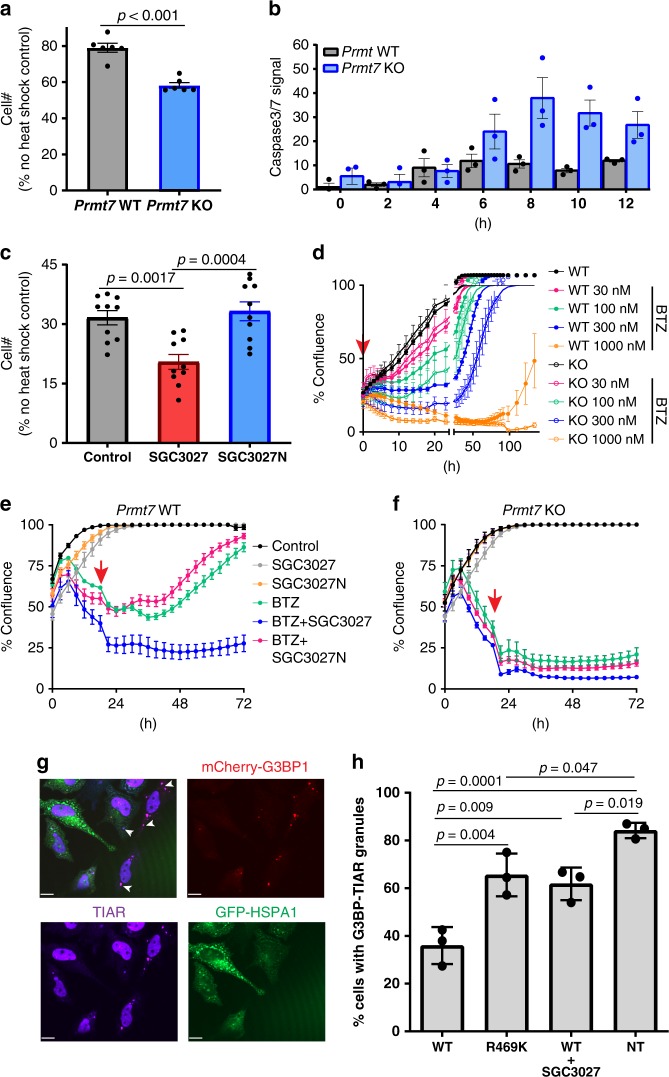
Protein arginine methyltransferases (PRMTs) regulate diverse biological processes and are increasingly being recognized for their potential as drug targets. Here we report the discovery of a potent, selective, and cell-active chemical probe for PRMT7. SGC3027 is a cell permeable prodrug, which in cells is converted to SGC8158, a potent, SAM-competitive PRMT7 inhibitor. Inhibition or knockout of cellular PRMT7 results in drastically reduced levels of arginine monomethylated HSP70 family stress-associated proteins. Structural and biochemical analyses reveal that PRMT7-driven in vitro methylation of HSP70 at R469 requires an ATP-bound, open conformation of HSP70. In cells, SGC3027 inhibits methylation of both constitutive and inducible forms of HSP70, and leads to decreased tolerance for perturbations of proteostasis including heat shock and proteasome inhibitors. These results demonstrate a role for PRMT7 and arginine methylation in stress response.
Conflict of interest statement
Y.I., N.S., H.F., T.S., K.H., S.T., M.T., and H.N. are current or former employees of Takeda. M.M.S., S.O., F.L., L.H., S.A., M.E., D.D., R.H., C.C.S., C.H.A., M.V., P.J.B., and D.B.-L. are current or former employees of the University of Toronto. The Structural Genomics Consortium is funded in part by Takeda. All other authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
