

Two Distinct Types of Eye-Head Coupling in Freely Moving Mice
- PMID: 32413309
- PMCID: PMC7284311
- DOI: 10.1016/j.cub.2020.04.042
Two Distinct Types of Eye-Head Coupling in Freely Moving Mice
Abstract
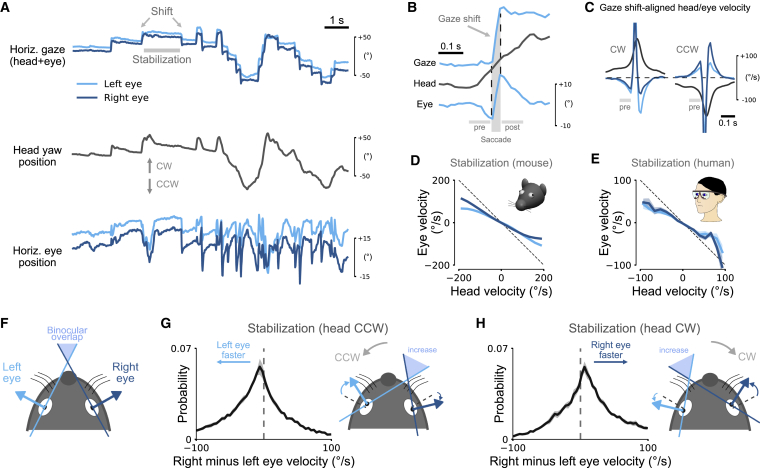
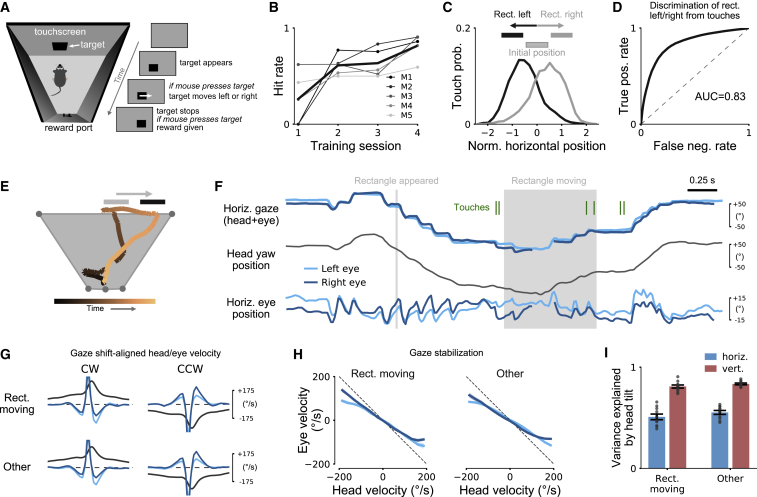
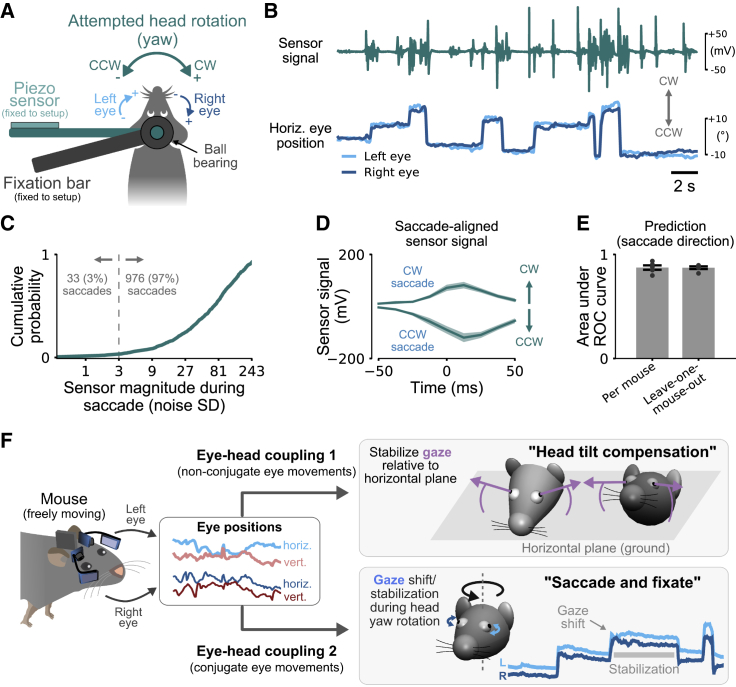
Animals actively interact with their environment to gather sensory information. There is conflicting evidence about how mice use vision to sample their environment. During head restraint, mice make rapid eye movements coupled between the eyes, similar to conjugate saccadic eye movements in humans. However, when mice are free to move their heads, eye movements are more complex and often non-conjugate, with the eyes moving in opposite directions. We combined head and eye tracking in freely moving mice and found both observations are explained by two eye-head coupling types, associated with vestibular mechanisms. The first type comprised non-conjugate eye movements, which compensate for head tilt changes to maintain a similar visual field relative to the horizontal ground plane. The second type of eye movements was conjugate and coupled to head yaw rotation to produce a "saccade and fixate" gaze pattern. During head-initiated saccades, the eyes moved together in the head direction but during subsequent fixation moved in the opposite direction to the head to compensate for head rotation. This saccade and fixate pattern is similar to humans who use eye movements (with or without head movement) to rapidly shift gaze but in mice relies on combined head and eye movements. Both couplings were maintained during social interactions and visually guided object tracking. Even in head-restrained mice, eye movements were invariably associated with attempted head motion. Our results reveal that mice combine head and eye movements to sample their environment and highlight similarities and differences between eye movements in mice and humans.
Keywords: eye movement; gaze; head movement; natural behavior; oculomotor system; pupil; vestibular system; vision.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
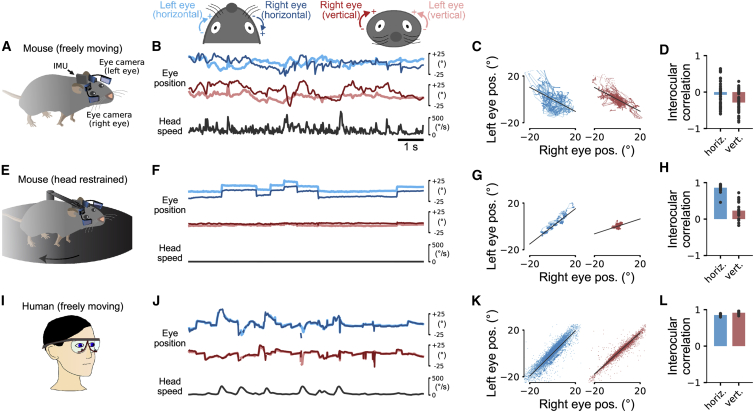
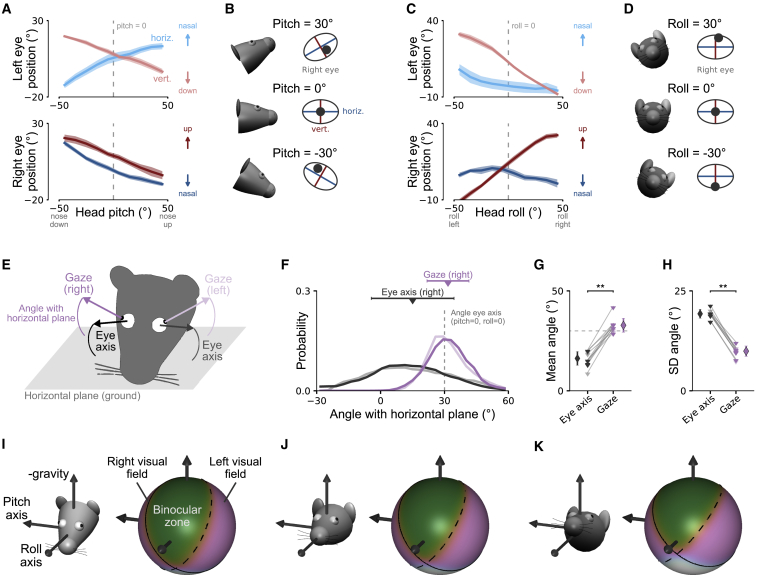
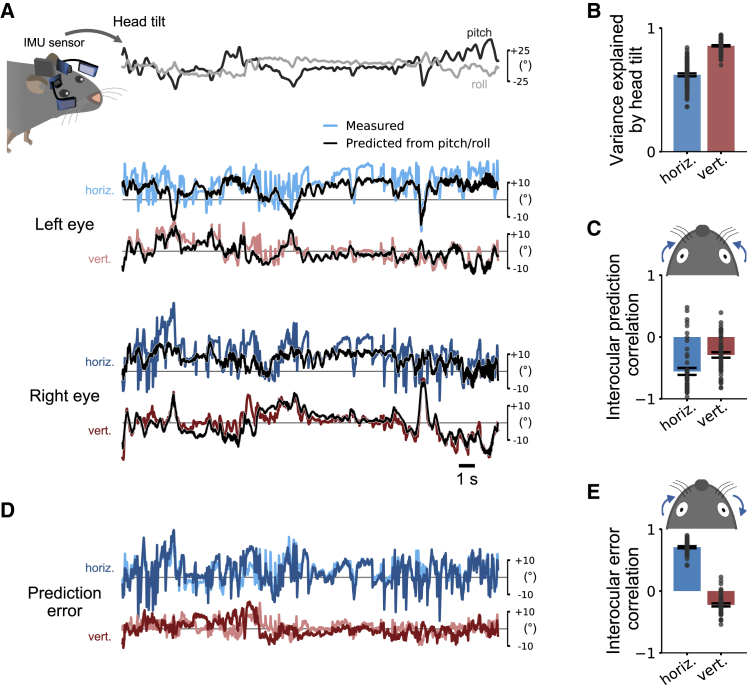
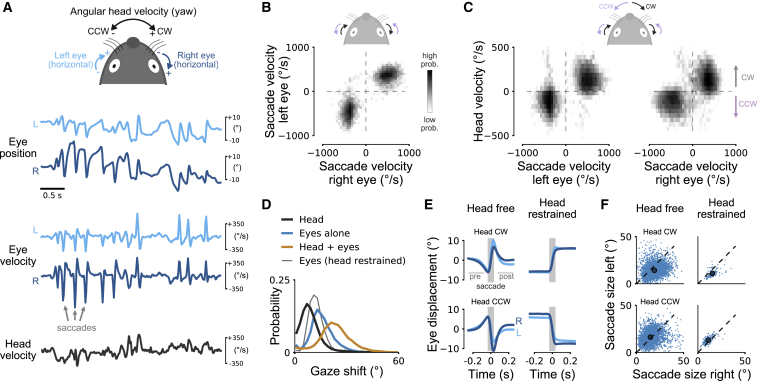
Comment in
-
Vision: How Mice Control Their View.Curr Biol. 2020 Jun 8;30(11):R635-R637. doi: 10.1016/j.cub.2020.04.063. Curr Biol. 2020. PMID: 32516609
References
-
- Kleinfeld D., Ahissar E., Diamond M.E. Active sensation: insights from the rodent vibrissa sensorimotor system. Curr. Opin. Neurobiol. 2006;16:435–444. - PubMed
-
- Gottlieb J., Oudeyer P.Y. Towards a neuroscience of active sampling and curiosity. Nat. Rev. Neurosci. 2018;19:758–770. - PubMed
-
- Land M., Mennie N., Rusted J. The roles of vision and eye movements in the control of activities of daily living. Perception. 1999;28:1311–1328. - PubMed
-
- Hayhoe M. Vision using routines: a functional account of vision. Vis. Cogn. 2000;7:43–64.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
