

Folding the Mitochondrial UPR into the Integrated Stress Response
- PMID: 32413314
- PMCID: PMC7230072
- DOI: 10.1016/j.tcb.2020.03.001
Folding the Mitochondrial UPR into the Integrated Stress Response
Abstract
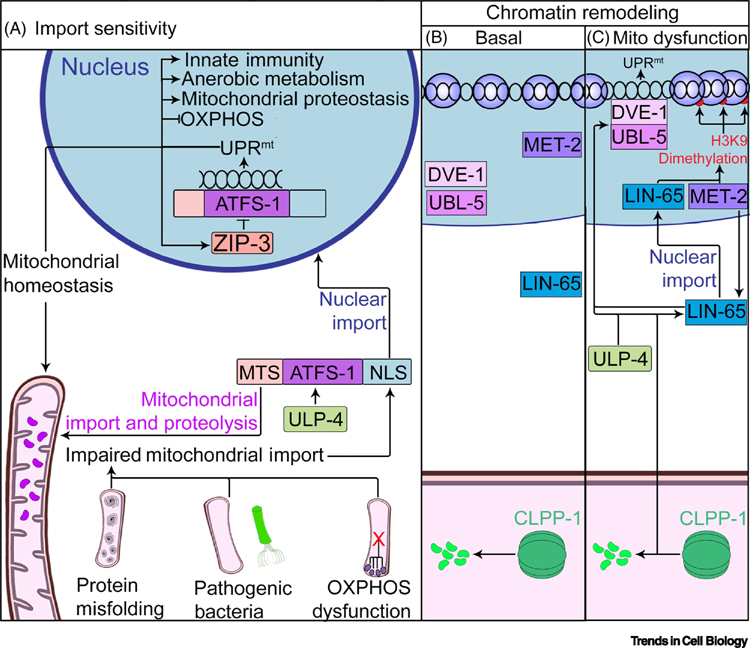
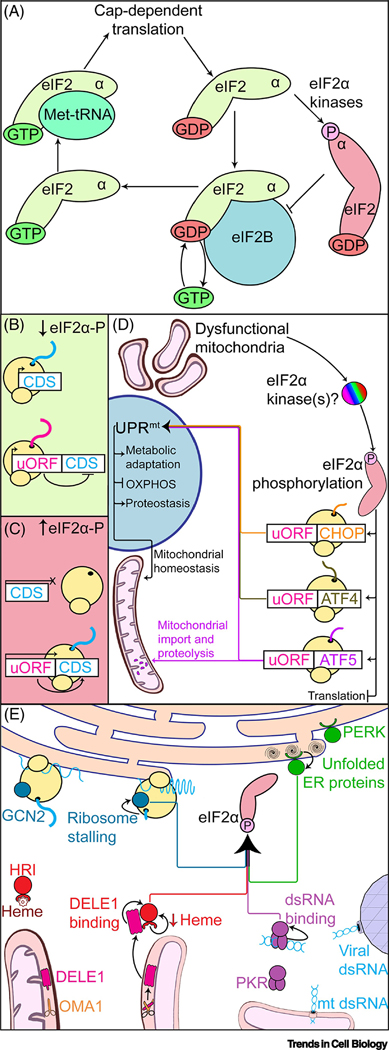
Eukaryotic cells must accurately monitor the integrity of the mitochondrial network to overcome environmental insults and respond to physiological cues. The mitochondrial unfolded protein response (UPRmt) is a mitochondrial-to-nuclear signaling pathway that maintains mitochondrial proteostasis, mediates signaling between tissues, and regulates organismal aging. Aberrant UPRmt signaling is associated with a wide spectrum of disorders, including congenital diseases as well as cancers and neurodegenerative diseases. Here, we review recent research into the mechanisms underlying UPRmt signaling in Caenorhabditis elegans and discuss emerging connections between the UPRmt signaling and a translational regulation program called the 'integrated stress response'. Further study of the UPRmt will potentially enable development of new therapeutic strategies for inherited metabolic disorders and diseases of aging.
Keywords: integrated stress response; mitochondria; mitochondrial unfolded protein response; stress signaling.
Copyright © 2020 Elsevier Ltd. All rights reserved.
Figures
References
-
- Chandel NS (2015). Evolution of Mitochondria as Signaling Organelles. Cell Metab 22, 204–206. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
