

Untangling Determinants of Enhanced Health and Lifespan through a Multi-omics Approach in Mice
- PMID: 32413334
- PMCID: PMC8214079
- DOI: 10.1016/j.cmet.2020.04.018
Untangling Determinants of Enhanced Health and Lifespan through a Multi-omics Approach in Mice
Abstract
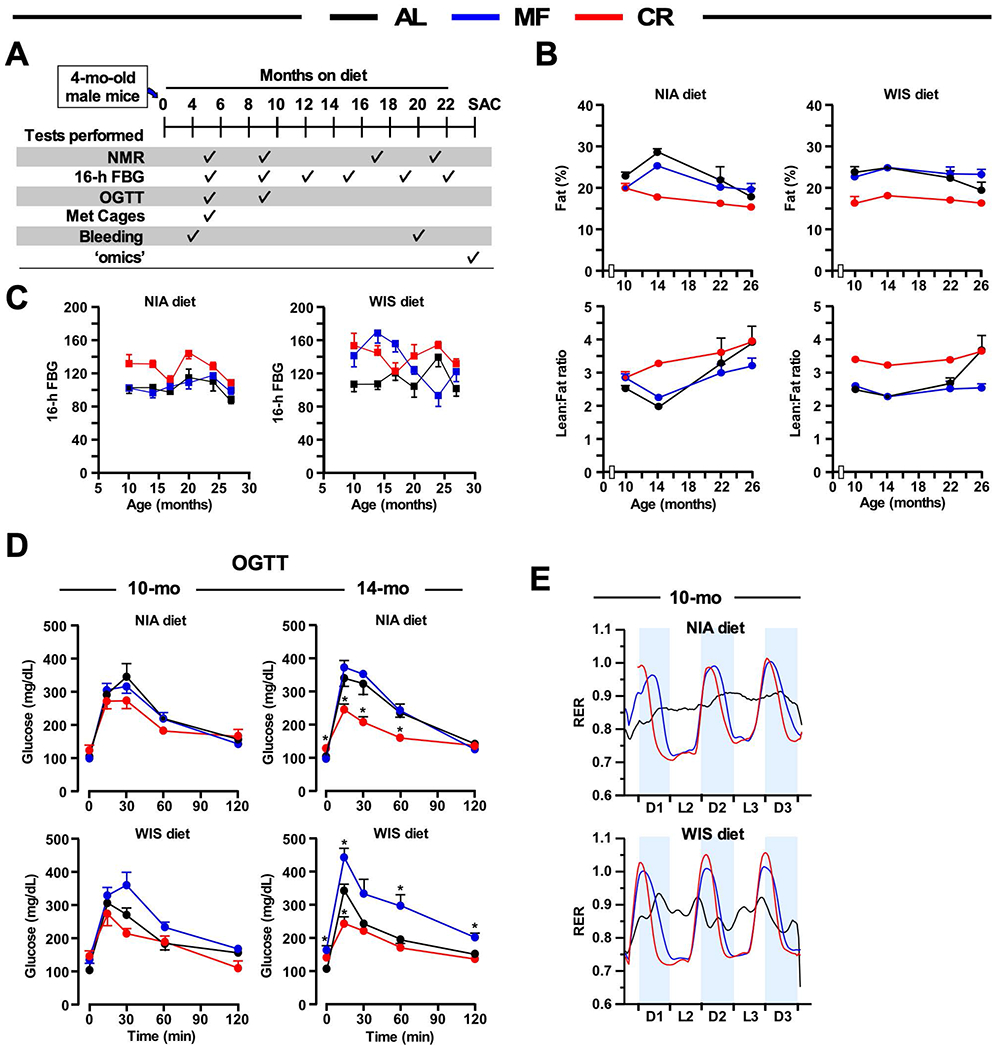
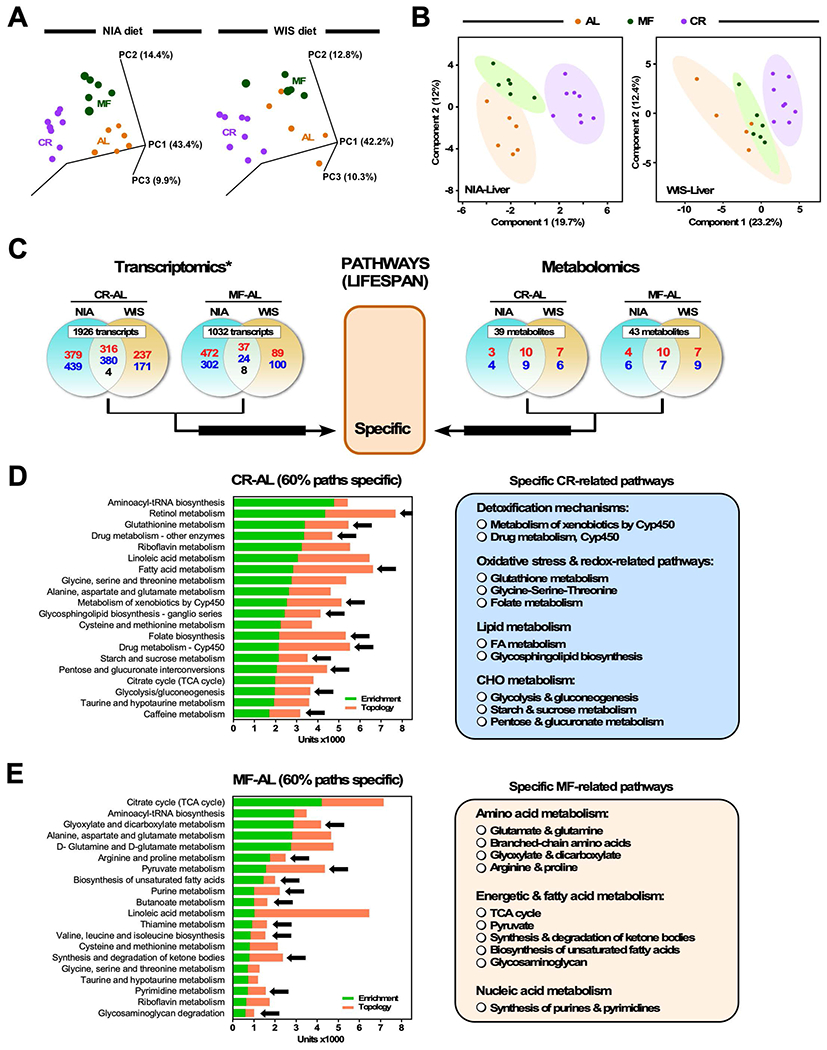
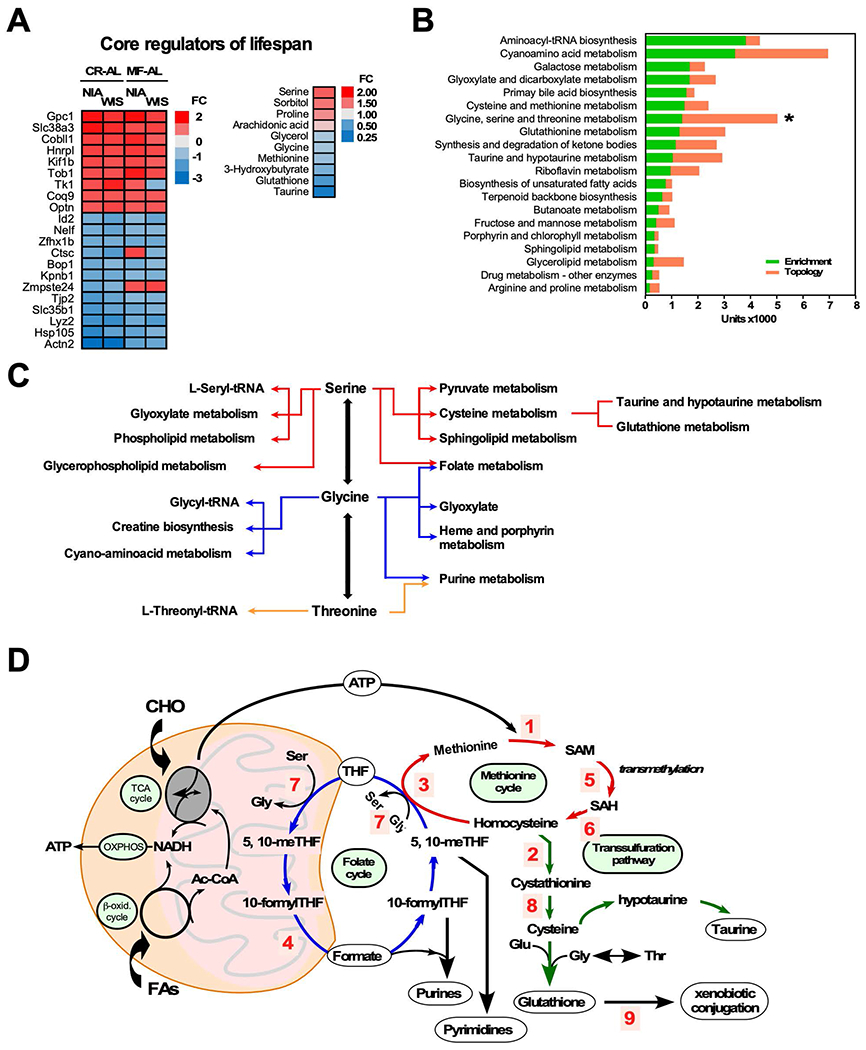
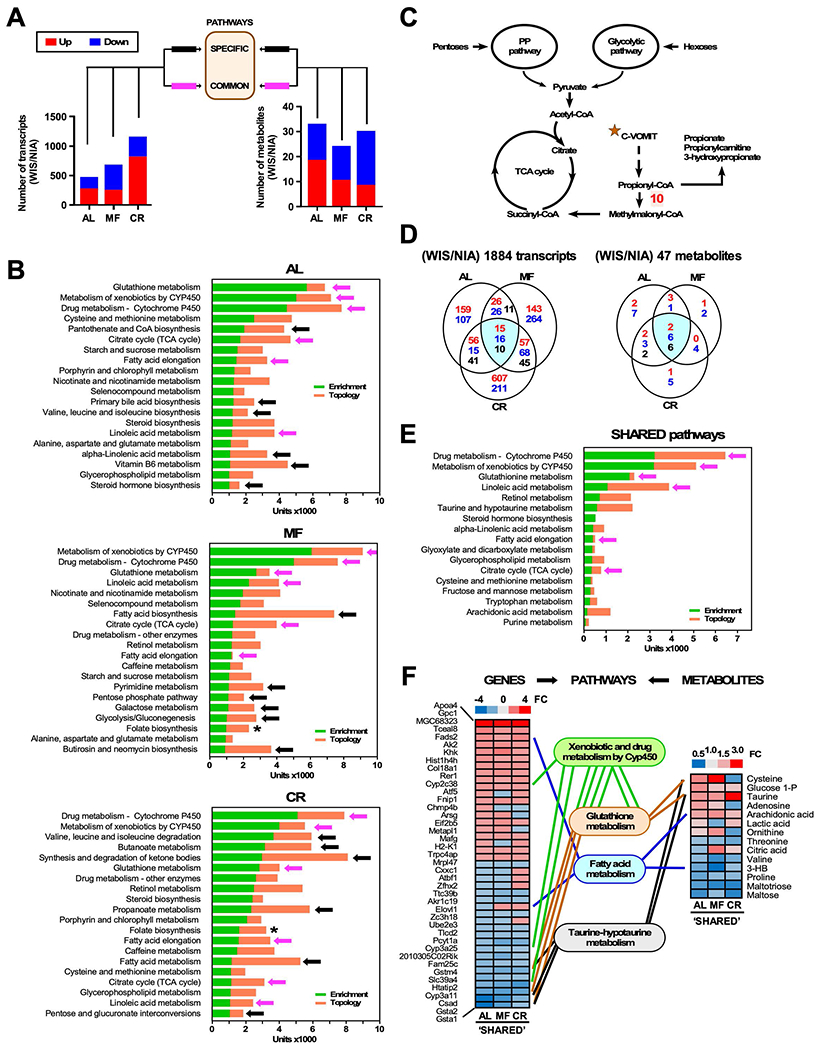
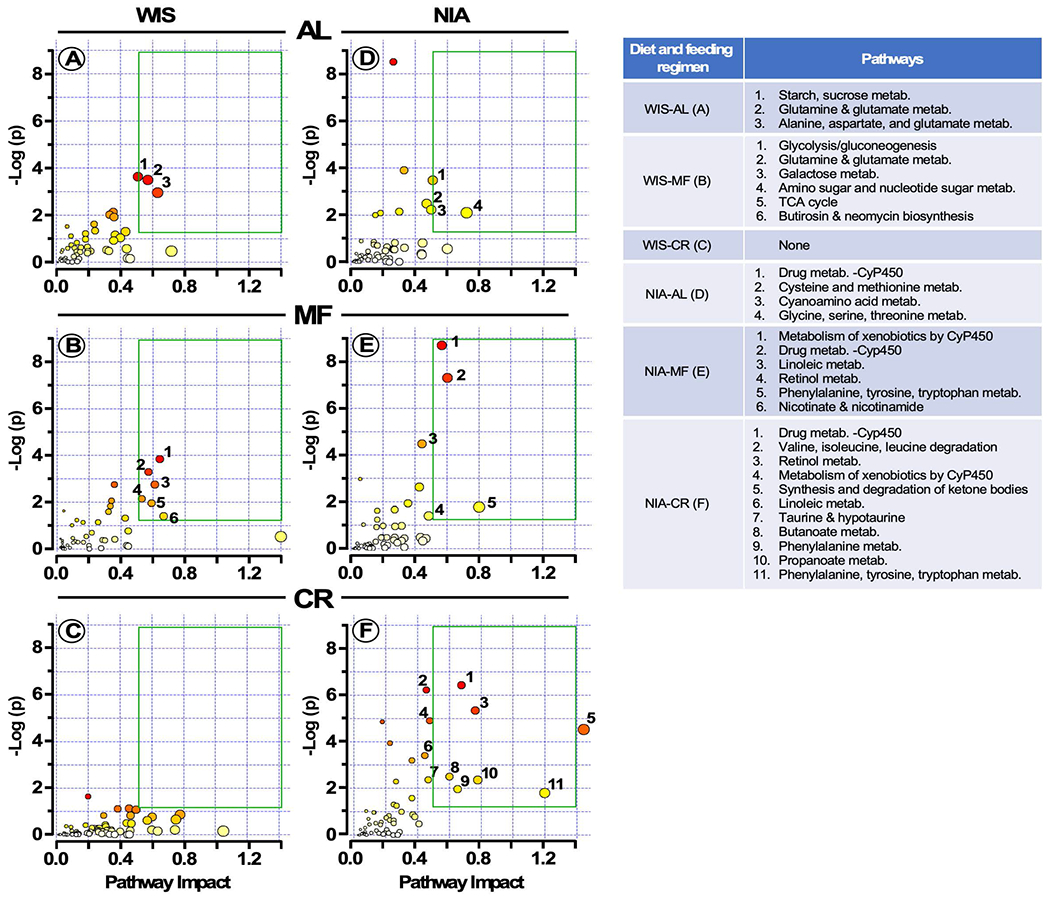
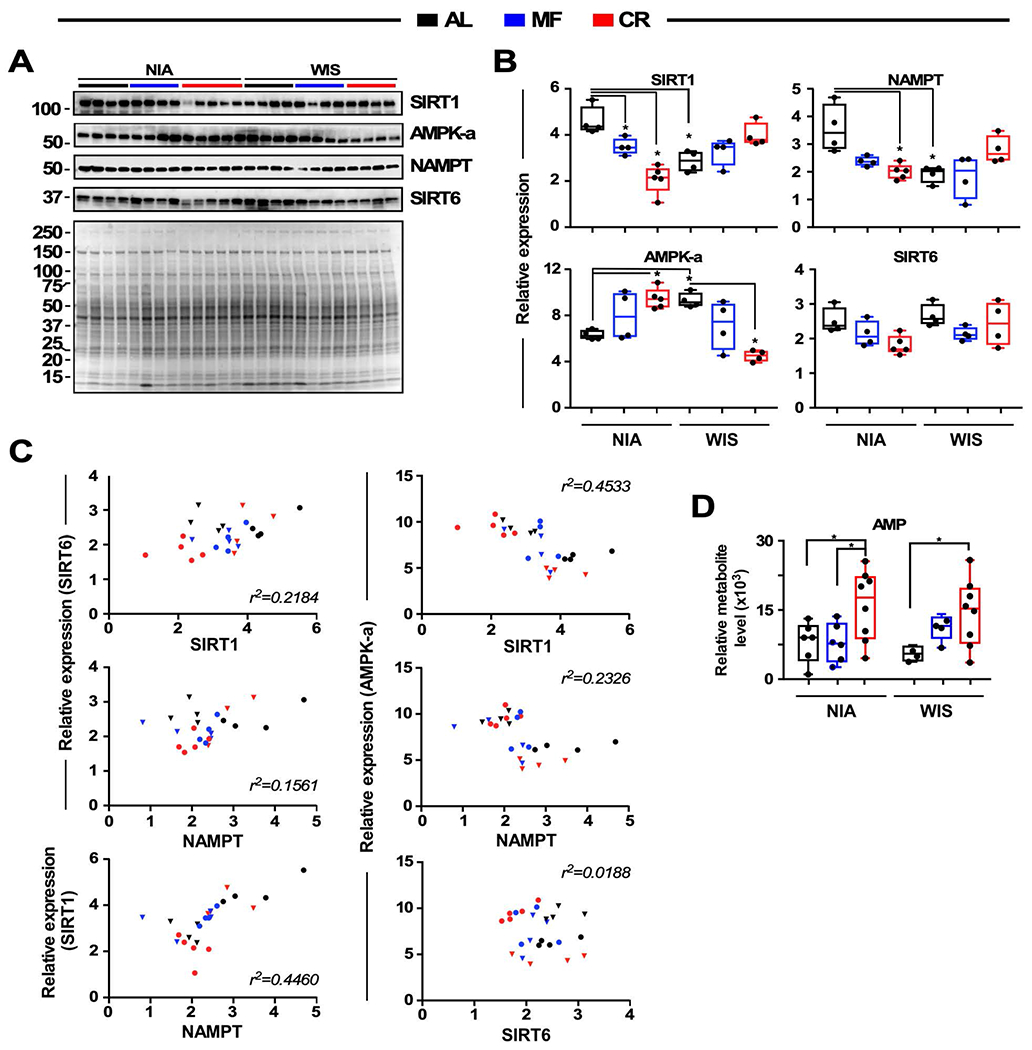
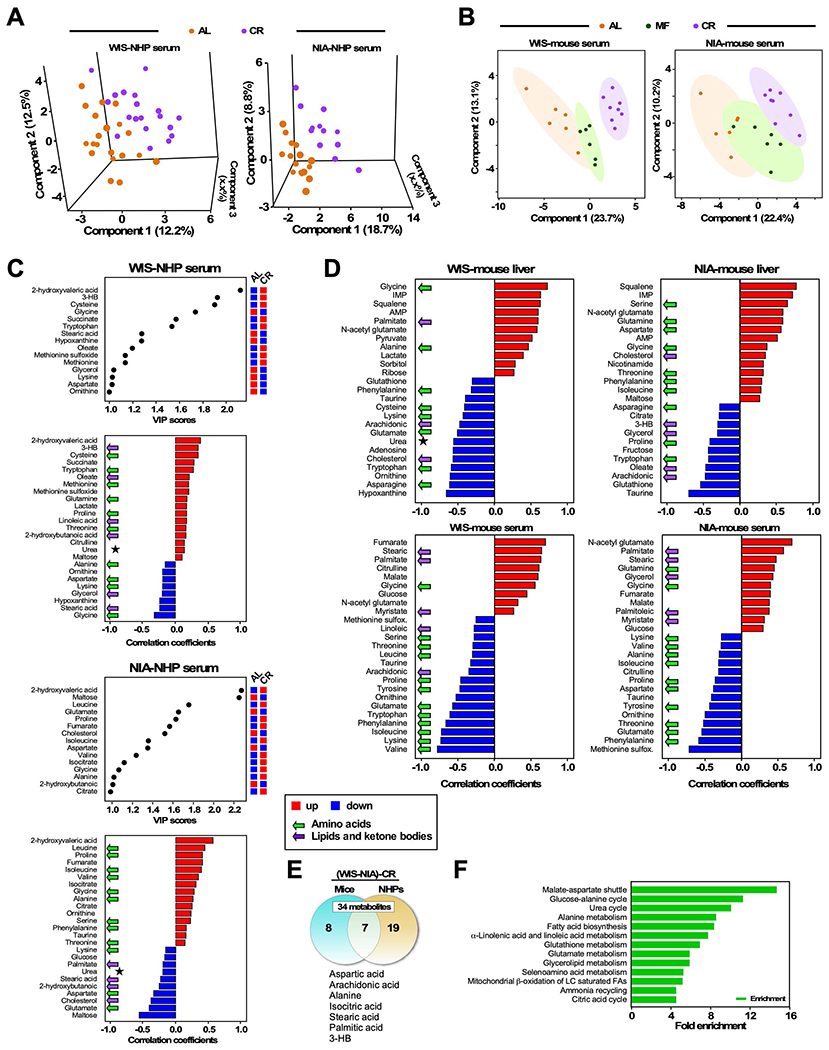
The impact of chronic caloric restriction (CR) on health and survival is complex with poorly understood underlying molecular mechanisms. A recent study in mice addressing the diets used in nonhuman primate CR studies found that while diet composition did not impact longevity, fasting time and total calorie intake were determinant for increased survival. Here, integrated analysis of physiological and multi-omics data from ad libitum, meal-fed, or CR animals was used to gain insight into pathways associated with improved health and survival. We identified a potential involvement of the glycine-serine-threonine metabolic axis in longevity and related molecular mechanisms. Direct comparison of the different feeding strategies unveiled a pattern of shared pathways of improved health that included short-chain fatty acids and essential PUFA metabolism. These findings were recapitulated in the serum metabolome from nonhuman primates. We propose that the pathways identified might be targeted for their potential role in healthy aging.
Keywords: aging; calorie restriction; calories; dietary interventions; dietary restriction; fasting; meal fed; metabolism; metabolomics; time-restricted feeding.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
References
-
- Aitken SM, and Kirsch JF, (2005). The enzymology of cystathionine biosynthesis: strategies for the control of substrate and reaction specificity. Arch Biochem Biophys 433, 166–175. - PubMed
-
- Al-Lahham SH, Peppelenbosch MP, Roelofsen H, Vonk RJ, and Venema K, (2010). Biological effects of propionic acid in humans; metabolism, potential applications and underlying mechanisms. Biochim Biophys Acta 1801, 1175–1183. - PubMed
-
- Baker EJ, Miles EA, Burdge GC, Yaqoob P, and Calder PC, (2016). Metabolism and functional effects of plant-derived omega-3 fatty acids in humans. Prog Lipid Res 64, 30–56. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
