

Kinetic diversity of amyloid oligomers
- PMID: 32414930
- PMCID: PMC7275774
- DOI: 10.1073/pnas.1922267117
Kinetic diversity of amyloid oligomers
Abstract
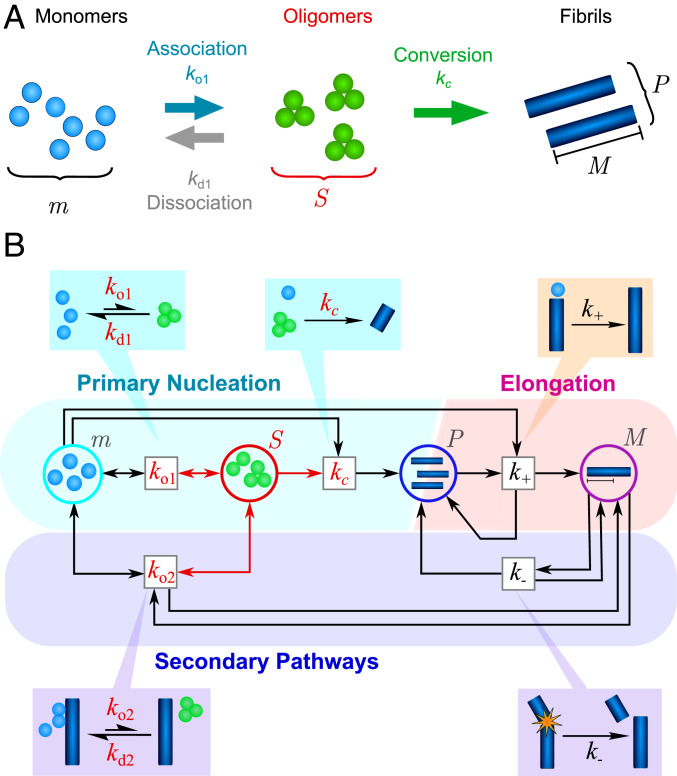
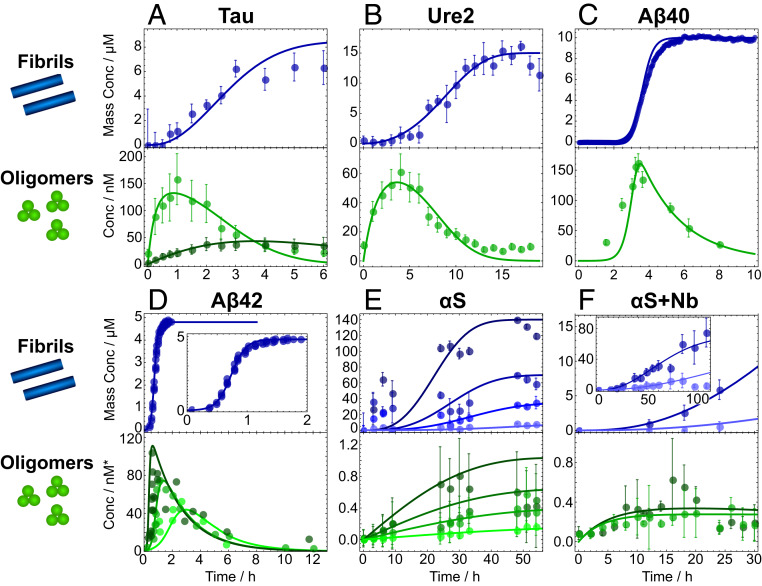
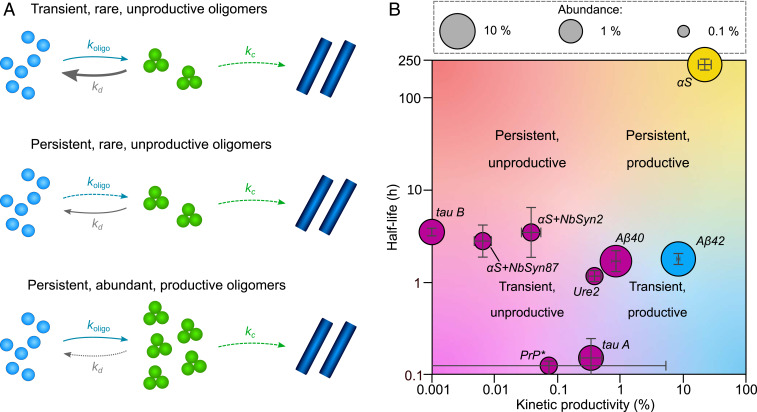
The spontaneous assembly of proteins into amyloid fibrils is a phenomenon central to many increasingly common and currently incurable human disorders, including Alzheimer's and Parkinson's diseases. Oligomeric species form transiently during this process and not only act as essential intermediates in the assembly of new filaments but also represent major pathogenic agents in these diseases. While amyloid fibrils possess a common, defining set of physicochemical features, oligomers, by contrast, appear much more diverse, and their commonalities and differences have hitherto remained largely unexplored. Here, we use the framework of chemical kinetics to investigate their dynamical properties. By fitting experimental data for several unrelated amyloidogenic systems to newly derived mechanistic models, we find that oligomers present with a remarkably wide range of kinetic and thermodynamic stabilities but that they possess two properties that are generic: they are overwhelmingly nonfibrillar, and they predominantly dissociate back to monomers rather than maturing into fibrillar species. These discoveries change our understanding of the relationship between amyloid oligomers and amyloid fibrils and have important implications for the nature of their cellular toxicity.
Keywords: Alzheimer’s; amyloid; kinetics; modeling; oligomers.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Lashuel H. A., Hartley D., Petre B. M., Walz T., Lansbury P. T., Neurodegenerative disease: Amyloid pores from pathogenic mutations. Nature 418, 291 (2002). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
