

Staufen 1 amplifies proapoptotic activation of the unfolded protein response
- PMID: 32415281
- PMCID: PMC7492261
- DOI: 10.1038/s41418-020-0553-9
Staufen 1 amplifies proapoptotic activation of the unfolded protein response
Erratum in
-
Correction: Staufen 1 amplifies proapoptotic activation of the unfolded protein response.Cell Death Differ. 2021 Dec;28(12):3374. doi: 10.1038/s41418-021-00734-x. Cell Death Differ. 2021. PMID: 33469232 Free PMC article. No abstract available.
Abstract
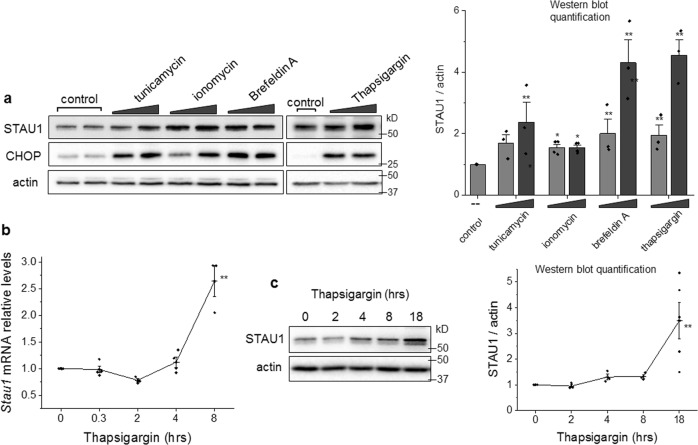
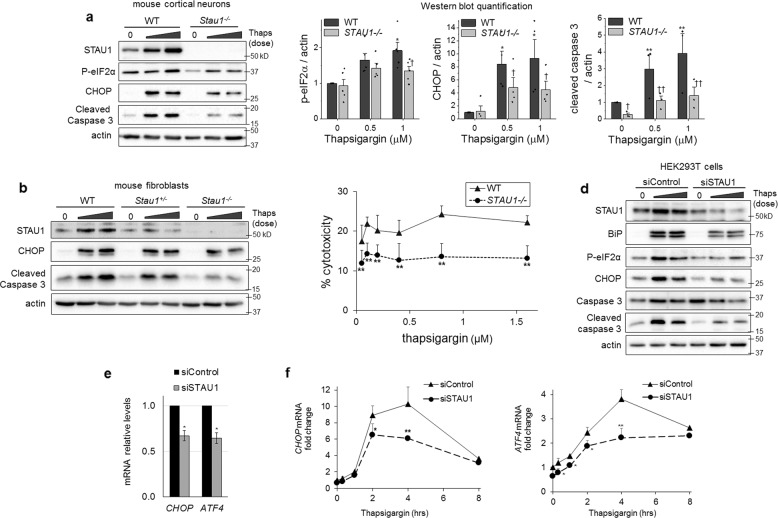
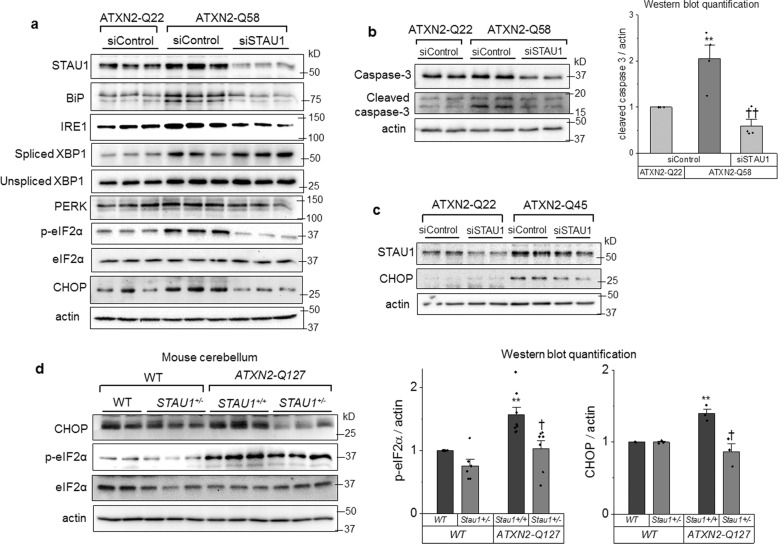
Staufen-1 (STAU1) is an RNA-binding protein that becomes highly overabundant in numerous neurodegenerative disease models, including those carrying mutations in presenilin1 (PSEN1), microtubule-associated protein tau (MAPT), huntingtin (HTT), TAR DNA-binding protein-43 gene (TARDBP), or C9orf72. We previously reported that elevations in STAU1 determine autophagy defects and its knockdown is protective in models of several neurodegenerative diseases. Additional functional consequences of STAU1 overabundance, however, have not been investigated. We studied the role of STAU1 in the chronic activation of the unfolded protein response (UPR), a common feature among neurodegenerative diseases and often directly associated with neuronal death. Here we report that STAU1 is a novel modulator of the UPR, and is required for apoptosis induced by activation of the PERK-CHOP pathway. STAU1 levels increased in response to multiple endoplasmic reticulum (ER) stressors, and exogenous expression of STAU1 was sufficient to cause apoptosis through the PERK-CHOP pathway of the UPR. Cortical neurons and skin fibroblasts derived from Stau1-/- mice showed reduced UPR and apoptosis when challenged with thapsigargin. In fibroblasts from individuals with SCA2 or with ALS-causing TDP-43 and C9ORF72 mutations, we found highly increased STAU1 and CHOP levels in basal conditions, and STAU1 knockdown restored CHOP levels to normal. Taken together, these results show that STAU1 overabundance reduces cellular resistance to ER stress and precipitates apoptosis.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures


References
-
- Paul S, Dansithong W, Gandelman M, Zu T, Ranum LPW, Figueroa KP, et al. Staufen blocks autophagy in neurodegeneration. 10.1101/659649. 2019.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
