

Simvastatin ameliorates altered mechanotransduction in uterine leiomyoma cells
- PMID: 32417359
- PMCID: PMC7609626
- DOI: 10.1016/j.ajog.2020.05.012
Simvastatin ameliorates altered mechanotransduction in uterine leiomyoma cells
Abstract
Background: Uterine leiomyomas, the most common tumors of the female reproductive system, are characterized by excessive deposition of disordered stiff extracellular matrix and fundamental alteration in the mechanical signaling pathways. Specifically, these alterations affect the normal dynamic state of responsiveness to mechanical cues in the extracellular environment. These mechanical cues are converted through integrins, cell membrane receptors, to biochemical signals including cytoskeletal signaling pathways to maintain mechanical homeostasis. Leiomyoma cells overexpress β1 integrin and other downstream mechanical signaling proteins. We previously reported that simvastatin, an antihyperlipidemic drug, has antileiomyoma effects through cellular, animal model, and epidemiologic studies.
Objective: This study aimed to examine the hypothesis that simvastatin might influence altered mechanotransduction in leiomyoma cells.
Study design: This is a laboratory-based experimental study. Primary leiomyoma cells were isolated from 5 patients who underwent hysterectomy at the Department of Gynecology and Obstetrics of the Johns Hopkins University Hospital. Primary and immortalized human uterine leiomyoma cells were treated with simvastatin at increasing concentrations (0.001, 0.01, 0.1, and 1 μM, or control) for 48 hours. Protein and mRNA levels of β1 integrin and extracellular matrix components involved in mechanical signaling were quantified by quantitative real-time polymerase chain reaction, western blotting, and immunofluorescence. In addition, we examined the effect of simvastatin on the activity of Ras homolog family member A using pull-down assay and gel contraction.
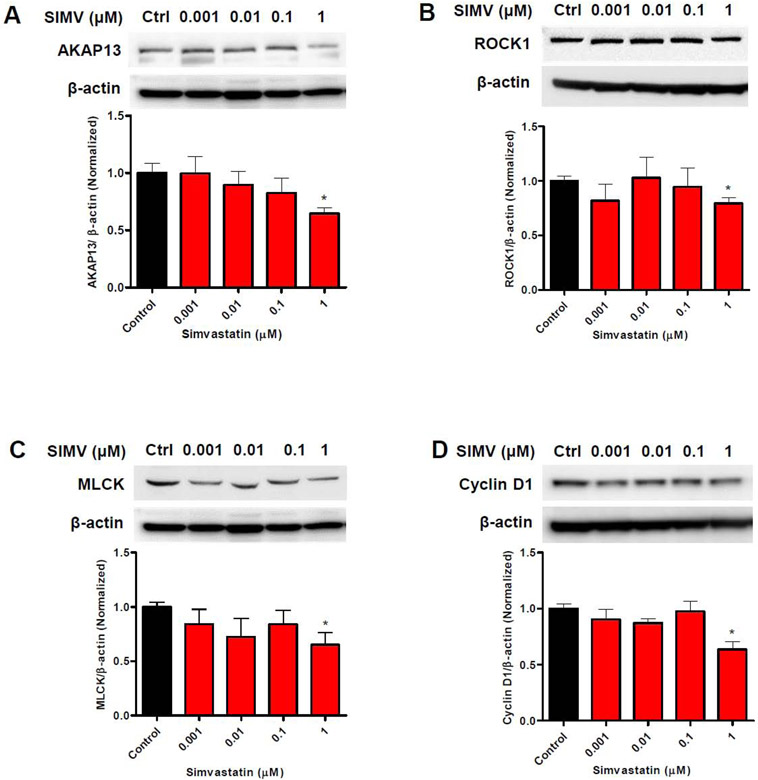
Results: We found that simvastatin significantly reduced the protein expression of β1 integrin by 44% and type I collagen by 60% compared with untreated leiomyoma cells. Simvastatin-treated cells reduced phosphorylation of focal adhesion kinase down to 26%-60% of control, whereas it increased total focal adhesion kinase protein expression. Using a Ras homolog family member A pull-down activation assay, we observed reduced levels of active Ras homolog family member A in simvastatin-treated cells by 45%-85% compared with control. Consistent with impaired Ras homolog family member A activation, simvastatin treatment reduced tumor gel contraction where gel area was 122%-153% larger than control. Furthermore, simvastatin treatment led to reduced levels of mechanical signaling proteins involved in β1 integrin downstream signaling, such as A-kinase anchor protein 13, Rho-associated protein kinase 1, myosin light-chain kinase, and cyclin D1.
Conclusion: The results of this study suggest a possible therapeutic role of simvastatin in restoring the altered state of mechanotransduction signaling in leiomyoma. Collectively, these findings are aligned with previous epidemiologic studies and other reports and support the need for clinical trials.
Keywords: AKAP13; FAK; MLCK; ROCK1; activated RhoA; cyclin D1; extracellular matrix; leiomyoma; mechanotransduction; simvastatin; type I collagen; β1 integrin.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest
William H. Catherino declares the following conflict of interest: Abbvie (Consultant); Allergan, now owned by Abbvie (Consultant); American Board of Obstetrics and Gynecology (Boards Examiner, Subject Matter Expert); American Society for Reproductive Medicine (Subject Matter Expert); Bayer (Consultant); EMD Serono (Consultant, wife is Senior Medical Director); Myovant (Consultant). Mostafa A. Borahay serves as Advisory Board member for Myovant Sciences.
Other authors report no conflict of interest
Figures










References
-
- Islam MS, Protic O, Stortoni P, et al. Complex networks of multiple factors in the pathogenesis of uterine leiomyoma. Fertil Steril 2013;100:178–93. - PubMed
-
- Bulun SE. Uterine fibroids. The New England journal of medicine 2013;369:1344–55. - PubMed
-
- Islam MS, Ciavattini A, Petraglia F, Castellucci M, Ciarmela P. Extracellular matrix in uterine leiomyoma pathogenesis: a potential target for future therapeutics. Hum Reprod Update 2018;24:59–85. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
